Nºmero 2, marzo
de 1999 ARACNET |

|
Apuntes sobre Biodiversidad y Conservaciµn de Insectos: Dilemas, Ficciones y ¢Soluciones? | Versiµn
electrµnica del artÚculo publicado en Bol.SEA, n¤ 20, monogrÃfico 'Los Artrµpodos y el Hombre' (1997, 468 pp.): 25-55, Sociedad Entomolµgica Aragonesa, Zaragoza (Espaþa) |
FermÚn MARTëN PIERA
e-mail: fermin@mncn.csic.es
Museo Nacional de Ciencias Naturales (C.S.I.C.).
Dpto. Biodiversidad y B. Evolutiva.
c/ JosÕ GutiÕrrez Abascal 2; 28006-Madrid.
Resumen
Introducciµn
1. La Biodiversidad
2. Medidas de Diversidad
3. Las causas de la Biodiversidad
4. El Paradigma de la Biodiversidad: Los Insectos
5. Conclusiones
6. Una ºltima reflexiµn sobre los insectos
Referencias
Tras un breve repaso conceptual sobre el significado del tÕrmino 'Biodiversidad', se revisan algunos aspectos teµricos y metodolµgicos relativos a la estimaciµn y medida de la diversidad biolµgica desde una triple perspectiva: ecolµgica, filogenÕtica y biogeogrÃfica. Seguidamente, se analizan algunos factores ecolµgicos e histµricos que determinan la Biodiversidad y se discute la importancia de los mÕtodos que combinan la informaciµn filogenÕtica y biogeogrÃfica, como criterios complementarios de las aproximaciones ecolµgicas tradicionales, a la hora de diseþar estrategias de conservaciµn fundamentadas en bases cientÚficas. En la ºltima parte del trabajo, se revisan algunos problemas inherentes a la conservaciµn de los insectos. A pesar de los innumerables 'servicios' ecolµgicos que nos prestan estos organismos, muchas de sus caracterÚsticas biolµgicas (dinÃmicas metapoblaciones, estacionalidad, variaciones demogrÃficas interanuales, ontogenia y ciclos vitales, adaptaciones ambientales, patrones de comportamiento reproductivos, etc.), invalidan los criterios y procedimientos de conservaciµn que se aplican usualmente a otros organismos. Se argumenta que gran parte de los 'dilemas' planteados por los Insectos, tienen su origen en una severa falta de conocimientos que se va acentuando por el creciente abandono de los estudios entomolµgicos. El nºmero de especialistas en nuestro paÚs es claramente insuficiente en relaciµn a la diversidad entomolµgica de nuestro territorio. Se estima que el nºmero actual de entomµlogos, habrÚa de incrementarse en un factor de 1,67.
Las lÚneas que siguen a continuaciµn constituyen una recopilaciµn y ampliaciµn de las ideas expuestas en diferentes conferencias, cursos y seminarios en los que he participado durante los ºltimos tres aþos. El mensaje que he transmitido en todos estos foros y que desarrollarÕ a continuaciµn, es la falta de conocimientos suficientes y rigurosos con los que, generalmente, se afrontan los problemas que plantea la medida, estimaciµn y conservaciµn de la diversidad biolµgica y, en particular, la del grupo de organismos paradigmÃtico de nuestro desconocimiento de la Biota terrestre: Los Insectos.
Hay tres razones por las que los biµlogos estamos interesados en la diversidad y su medida:
1) La diversidad es un concepto intelectualmente muy atractivo que sigue suscitando un considerable debate. A escala ecolµgica, los bien conocidos patrones de variaciµn espacio-temporal de la diversidad, continºan estimulando la mente de todos cuantos nos dedicamos al estudio de la diversidad biolµgica, desde cualquier enfoque.
2) Frecuentemente se acostumbra a pensar que las medidas de diversidad son buenos indicadores del estado de ‘salud’ del ecosistema.
3) La diversidad es uno de los temas centrales de la BiologÚa. A escala evolutiva, la explicaciµn de la diversidad constituye la cuestiµn esencial que intenta explicar el paradigma darwiniano de evoluciµn por selecciµn natural.
Independientemente de la problemÃtica cientÚfica y/o conservacionista que plantea el estudio de la diversidad biolµgica, la diversidad entomolµgica complica aºn mÃs muchos de estos problemas en especial, desde el punto de vista metodolµgico. El estudio de los insectos evidencia, ademÃs, muchas ficciones conservacionistas y no parece aportar soluciones, al menos a corto plazo, dado que los insectos, con demasiada frecuencia, quedan excluidos de muchos de los estudios sobre Conservaciµn de êreas y Organismos. Sin embargo, no podemos olvidar que el Hombre està daþando la estructura de la diversidad en la Biosfera y los insectos son el componente mayoritario de esta Biodiversidad. Por ello, desde una perspectiva cientÚfica no se entiende bien que se excluya a los insectos de los anÃlisis sobre el reparto geogrÃfico, evaluaciµn y conservaciµn de la biodiversidad ya que, generalmente, constituyen la fracciµn mÃs importante de la diversidad de un territorio.
El trabajo està estructurado en dos grandes bloques. En el primero se revisan algunos aspectos teµricos relativos a la Biodiversidad y su medida a travÕs de diferentes aproximaciones ecolµgicas, taxonµmicas y corolµgicas (Secciones 1 y 2) y se hace un breve repaso de los factores causales o generadores de diversidad (Secciµn 3). El segundo bloque, està dedicado a la problemÃtica particular que plantea el estudio y conservaciµn de los insectos, de acuerdo con las caracterÚsticas singulares de sus poblaciones, distribuciµn, rareza, ciclos biolµgicos y megadiversidad (Secciµn 4).
Mucha gente tiene una idea intuitiva de la diversidad y no tiene ninguna dificultad en aceptar que, por ejemplo, el bosque lluvioso tropical aloja mayor nºmero de seres vivos que un desierto o que existe una alta diversidad de organismos en los arrecifes de coral. Pero ¢quÕ es la Biodiversidad?, ¢cµmo podemos definirla?. A menudo el uso y abuso de un vocablo termina vaciÃndolo de contenido. Algo parecido comienza a suceder con la Biodiversidad. No han faltado incluso detractores del tÕrmino. Argumentan que es tal la magnitud conceptual de este neologismo que expresa todo y, al mismo tiempo, nada. Algo hay de razµn en ello, pero sµlo algo. AsÚ, para Hurlbert (1971) la diversidad es un ‘non-concept’. Para otros es, simplemente, un pseudovocablo cuyos usuarios comparten una definiciµn intuitiva (Salt, 1979). Otros en cambio, algo mÃs moderados, aducen que al menos, no es fÃcil definirla. Ello se debe a que la diversidad en un contexto ecolµgico, engloba dos componentes: variedad y abundancia relativa de especies (Magurran, 1988).
En todo caso es innegable que en la ºltima dÕcada la palabra ‘Biodiversidad’ parece haber surgido mÃgicamente llenando el vocabulario de nuestra sociedad a todos los niveles: artÚculos cientÚficos, proyectos de investigaciµn, cumbres internacionales, medios de comunicaciµn, informes gubernamentales, etc. El uso creciente de este vocablo en la literatura cientÚfica, ha sido calificado incluso de dramÃtico (Harper & Hawksworth, 1996). Sin embargo, en Ciencia los conceptos son importantes. Por ello, parece razonable detenerse a reflexionar quÕ significa exactamente una palabra tan ampliamente usada en los ºltimos aþos y no solamente en foros cientÚficos. ¢Es una nueva botella para el vino aþejo de las viejas ideas como seþalan Harper & Hawksworth (1996) µ nos estamos refiriendo a una cuestiµn fundamental de la Ciencia?. MÃs aºn, ¢estamos asistiendo al nacimiento de una disciplina emergente, netamente multidisciplinar que denominamos Biodiversidad, o por el contrario, este vocablo es un neologismo vacÚo de contenido, la nueva etiqueta de otras disciplinas mÃs aþejas, con mÕtodos y objetivos propios (SistemÃtica, Filogenia, BiogeografÚa, EcologÚa), con la que acceder a nuevos recursos y seguir haciendo las mismas cosas? (di Castri, 1993). Ambos fenµmenos estÃn sucediendo simultÃneamente. Como suele suceder en la historia del pensamiento, los nuevos paradigmas conviven durante un tiempo con las viejas ideas.
Fueron Norse y colaboradores (1986) quienes primero generalizaron el tÕrmino Biodiversidad a tres niveles: diversidad genÕtica dentro de cada especie, diversidad de especies (riqueza µ nºmero de especies) y diversidad ecolµgica (comunidades). Pero la autÕntica explosiµn del tÕrmino, aconteciµ dos aþos mÃs tarde cuando Wilson (1988) se refiriµ a la Biodiversidad como el patrimonio o riqueza biµtica singular e irrepetible de cada lugar, regiµn µ continente y, en ºltima instancia, de toda la humanidad; el rÚo de la vida del que nos habla Margalef. Nuestro eminente ecµlogo hace ademÃs una distinciµn semÃntica fundamental. Para Margalef diversidad y Biodiversidad no son vocablos sinµnimos que expresen conceptos idÕnticos. La Biodiversidad es el ‘Gran Diccionario de la Vida’, el inmenso caudal de formas presentes y extintas en los que se manifiesta el devenir histµrico de la Vida. En cambio, la Diversidad es, en el sÚmil de Margalef, el ‘Lenguaje’ que representa los experimentos de evoluciµn que van enriqueciendo el Diccionario.
Una de las definiciones mÃs ampliamente difundidas es la de McNeely et al. (1990). Segºn este autor la Biodiversidad es un paraguas conceptual que engloba la variedad de la Naturaleza, incluyendo el nºmero y frecuencia de ecosistemas, especies y genes representados por un conjunto de organismos.
Ahora bien, a pesar de toda esta reciente efervescencia teµrica, cabe preguntarse si no estaremos poniendo de moda uno de los problemas mÃs clÃsicos y fundamentales de la BiologÚa, tal vez el problema central; me he referido a ello anteriormente en la Introducciµn (ver punto 3). En efecto, creo que la respuesta mÃs general y sencilla a la pregunta inicial (¢quÕ es la Biodiversidad?), no es otra que la que emana de la propia TeorÚa Evolutiva: La diversidad biolµgica es el resultado mÃs genuino del proceso evolutivo, que se manifiesta a todos los niveles jerÃrquicos de la vida: de las molÕculas a los ecosistemas, pasando por los genes, las cÕlulas, los individuos, las poblaciones y las comunidades (Solbrig, 1991 a y b). Es, como ya he repetido, el problema esencial que afronta Darwin (1859) en su famoso libro, ‘Del origen de las especies por selecciµn natural...’. Descendencia con variaciµn, mutaciµn y selecciµn son, en tÕrminos ortodoxos, las causas que determinan la cantidad de seres vivos que existen en un tiempo y espacio determinados. Esta perspectiva histµrica es la que manifiesta en toda su dimensiµn, la caracterÚstica esencial del estado singular de la materia que denominamos Vida: el hecho de ser diversa, variable y variada.
En todos los conceptos y definiciones de Biodiversidad, convergen dos dimensiones estrechamente relacionadas que dependen de la escala de enfoque y de la escala de anÃlisis (genes, individuos, poblaciones, especies, comunidades, ecosistemas): la histµrico-evolutiva y la ecolµgico-funcional. La primera se consagra al estudio de los Patrones estructurales y espacio-temporales en los que se expresan las relaciones genealµgicas de los organismos, ilustradas en las clasificaciones jerÃrquicas y la segunda, a los Procesos e interrelaciones funcionales que acontecen en la intimidad de los propios organismos y en el seno de los ecosistemas.
Si aceptamos que la diversidad es una
propiedad de los seres vivos y, por tanto, algo mÃs que el nºmero de especies en un
tiempo y lugar, hemos de plantearnos cµmo medirla. Es indudable que sin una
cuantificaciµn de la diversidad biolµgica no podremos movilizar una ciencia seria y
rigurosa de la Biodiversidad. AdemÃs, a menudo necesitamos diferentes aproximaciones
metodolµgicas (medidas) para resolver problemas de distinta Úndole. Por ejemplo, para
establecer una red de reservas naturales que alojen la mayor diversidad posible en el
mÚnimo nºmero de Ãreas protegidas, precisaremos medidas de diversidad espacial (![]() /Ô/
/Ô/![]() -diversidad) y
aproximaciones que combinen la informaciµn filogenÕtica y biogeogrÃfica. En cambio, si
nuestro objetivo es investigar la importancia funcional de una o varias especies en un
ecosistema degradado, en franco retroceso, con especies en peligro de extinciµn, nuestra
investigaciµn tendrà que dirigirse a conocer las relaciones funcionales de las especies
en el seno del ecosistema, averiguar cuÃl es el estado de los procesos ecolµgicos que
tienen lugar y si es posible la aplicaciµn de medidas que permitan la inversiµn del
proceso de regresiµn, hasta la restauraciµn de las interrelaciones funcionales
originales.
-diversidad) y
aproximaciones que combinen la informaciµn filogenÕtica y biogeogrÃfica. En cambio, si
nuestro objetivo es investigar la importancia funcional de una o varias especies en un
ecosistema degradado, en franco retroceso, con especies en peligro de extinciµn, nuestra
investigaciµn tendrà que dirigirse a conocer las relaciones funcionales de las especies
en el seno del ecosistema, averiguar cuÃl es el estado de los procesos ecolµgicos que
tienen lugar y si es posible la aplicaciµn de medidas que permitan la inversiµn del
proceso de regresiµn, hasta la restauraciµn de las interrelaciones funcionales
originales.
En todo caso, las dimensiones ecolµgica y evolutiva de la diversidad biolµgica proporcionan los marcos teµricos y metodolµgicos de referencia, en los que desarrollar medidas adecuadas para resolver las numerosas cuestiones que plantea en la actualidad, los problemas relativos a la conservaciµn y preservaciµn de la Biodiversidad. Aunque la EcologÚa es una ciencia biolµgica mÃs joven que las clÃsicas disciplinas evolutivas (SistemÃtica, Filogenia, BiogeografÚa), paradµjicamente, las medidas ecolµgicas tienen una larga tradiciµn histµrica, en comparaciµn con las medidas evolutivas, las cuales, apenas tienen diez aþos de desarrollo intelectual.
|
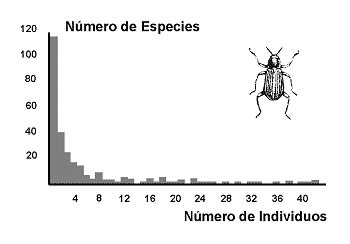
Figura 1. Distribuciµn de abundancia en una comunidad de coleµpteros del rÚo TÃmesis. Modificado de Magurran (1988). Datos de Williams (1964). |
2.1. Medidas Ecolµgicas de Diversidad
En el sentido ecolµgico mÃs estricto la diversidad - un concepto derivado de la TeorÚa de la Informaciµn - es una medida de la heterogeneidad del sistema, es decir, de la cantidad y proporciµn de los diferentes elementos que contiene. AdemÃs del significado que en sÚ misma tiene la diversidad, es tambiÕn un parÃmetro muy ºtil en el estudio, descripciµn y comparaciµn de las comunidades ecolµgicas. Dado que la diversidad en una comunidad es una expresiµn del reparto de recursos y energÚa, su estudio es una de las aproximaciones mÃs ºtiles en el anÃlisis comparado de las comunidades, o incluso de regiones naturales (Halffter & Ezcurra, 1992).
De todos los Úndices descritos en la literatura, los dos mÃs clÃsicos son el ëndice de Simpson y el ëndice de Shanon-Wienner. El primero es una medida de Dominancia y se expresa como:
![]() =
= ![]() pi2
pi2
siendo pi = ni /N, donde ni es el nºmero de individuos de la especie ‘i’ y N es la abundancia total de las especies. Con otras palabras, pi es la abundancia proporcional de la especie ‘i’.
A medida que el Úndice se incrementa,
la diversidad decrece. Por ello el ëndice de Simpson se presenta habitualmente como 1/![]() =
1/
=
1/![]() pi2 que expresa, en realidad, una medida de la
dominancia, como se acaba de indicar. Por tanto, el Úndice de Simpson sobrevalora las
especies mÃs abundantes en detrimento de la riqueza total de especies.
pi2 que expresa, en realidad, una medida de la
dominancia, como se acaba de indicar. Por tanto, el Úndice de Simpson sobrevalora las
especies mÃs abundantes en detrimento de la riqueza total de especies.
El ëndice de Shannon-Wienner, a veces incorrectamente denominado ëndice de Shannon-Weaver (Krebs, 1985), procede de la TeorÚa de la Informaciµn y se expresa como:
H’ = - ![]() pi
ln pi
pi
ln pi
Otro de los Úndices mÃs difundidos es el ëndice de Margalef (1958):
Dmg = (S-1)/ln N
|
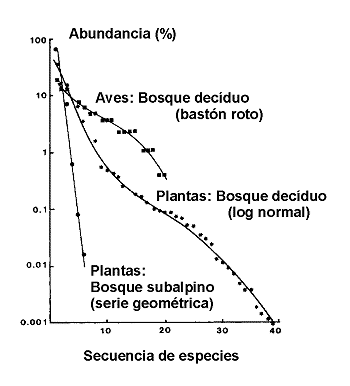
Figura 2. Curvas de abundancia de especies ilustrativas de tres modelos de distribuciµn: 'bastµn roto', log normal y serie geomÕtrica. Modificado de Magurran (1988). Datos de Whittaker (1970). |
Estos Úndices pueden derivarse de los denominados nºmeros de diversidad de Hill (Ludwig & Reynolds, 1988). El mÕtodo de Hill permite ordenar estos Úndices de acuerdo a su tendencia a sobrevalorar bien la riqueza de especies (dando mÃs ‘peso’ a las especies menos abundantes y poco comunes o raras), o bien, la dominancia, es decir, dando mayor Õnfasis a las especies mÃs abundantes. Hill (1973) propuso un mÕtodo elegante para describir las relaciones entre todos estos Úndices, definiendo un Úndice de diversidad como la abundancia proporcional media recÚproca y fue capaz de clasificarlos de acuerdo al peso que cada uno de ellos otorgaba a las especies mÃs raras:
Na = (p1a + p2a + p3a +....+ pna)1/(1-a)
Na Na es al a orden de diversidad y pn es la abundancia proporcional de la especie de orden n.
Si a = 0; N0 = S nºmero de especies totales en la comunidad.
Si a = 1; N1 = e H’
Si a = 2; N2 = 1/![]() = 1/ pi2.
= 1/ pi2.
Siempre serà cierto que N0 > N1 > N2.
En el primer caso todas las especies, incluso las raras, son igualmente pesadas. En el segundo se pesan las especies abundantes y, en el tercero, las especies muy abundantes.
El otro parÃmetro descriptivo de las relaciones de abundancia, es la Equitatividad. La equitatividad tambiÕn se puede medir de muchas formas. Una de las mÃs sencillas es estimarla a partir de la abundancia de la especie dominante:
E = 1/sp1
No obstante, una de las mÃs frecuentes es la razµn que expresa la equitatividad como la diversidad H’ (encontrada) con relaciµn al mÃximo valor que H’ puede alcanzar cuando todas las especies muestran idÕnticas abundancias; es la familiar ‘J’ de Pielou (1975, 1977):
E = H’/Hmax = H’/lnS
Para un desarrollo mÃs extenso de los indices de diversidad, dominancia y equitatividad, el lector interesado puede consultar cualquier texto de EcologÚa general. Por su claridad y amplitud resultan muy recomendables los de Magurran (1988) y Ludwig & Reynolds (1988). Son igualmente recomendables algunos artÚculos que tratan con profundidad diferentes aspectos y problemas relativos a la medida ecolµgica de la diversidad; por ejemplo, Alatalo (1981) en lo que se refiere a Úndices de equitatividad.
Por su mayor complejidad, no se trata aquÚ el tema de la estimaciµn de la riqueza de especies. No obstante, merece la pena citar tres artÚculos de sÚntesis que tratan en detalle, con claridad y rigor este interesante tema: Hetlshe & Forrester (1983), BaltanÃs (1991) y Colwell & Coddington (1994).
|
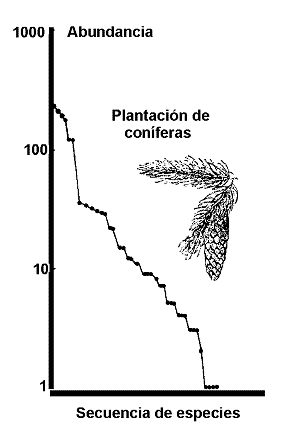
Figura 3. Curva de abundancia de especies en una plantaciµn de conÚferas de Irlanda. Modificado de Magurran (1988). |
2.1.1. Modelos de distribuciµn
Las especies se distribuyen, normalmente, segºn jerarquÚas de abundancia desde algunas especies muy abundantes hasta algunas muy raras. Generalmente, en las comunidades lo normal es que haya bastantes especies raras, pocas especies abundantes y muchas especies con una abundancia intermedia (Figura 1). Esta parece una regla que tambiÕn se cumple en las sociedades humanas: hay bastantes personas pobres, pocas muy ricas y mucha clase media. Es muy probable que este tipo de relaciµn especies/abundancia, se dÕ siempre que los elementos del sistema interaccionan y compiten por unos recursos limitados. Esta observaciµn condujo a los modelos de distribuciµn de abundancia de especies.
Se han descrito cuatro modelos principales: Las series logarÚtmicas (log-series) de Fischer, Corbet & Williams (1943), el modelo lognormal (Preston, 1948), las series geomÕtricas y el denominado modelo de ‘bastµn roto’ (MacArthur, 1957). Como se aprecia en la figura 2, hay una progresiµn desde el modelo geomÕtrico con unas pocas especies muy abundantes y las restantes muy raras y escasas a travÕs de las log-series y la distribuciµn log-normal, en las que las especies con abundancias intermedias son las mÃs comunes hasta el modelo en bastµn roto en el que casi todas las especies son igualmente abundantes. Estos cuatro modelos se han interpretado clÃsicamente en tÕrminos de distribuciµn de recursos, en donde la abundancia de una especie es, de alguna forma, equivalente a la porciµn de nicho ocupado por cada especie en la comunidad.
Me centrarÕ ºnicamente en las dos primeras por ser las que explican la mayor parte de los datos experimentales.
El modelo de series logarÚtmicas de Fisher, Corbet & Williams representµ el primer intento de describir matemÃticamente la relaciµn entre el nºmero de especies y el nºmero de individuos que representan a cada una de ellas. Williams, un lepidopterµlogo que trabajµ en el archipiÕlago malayo, planteµ a Fischer la siguiente cuestiµn: Conociendo cuantas especies se habÚan colectado y la abundancia de cada una de ellas y aceptando que la colecciµn de Williams no contuviera todas las mariposas ¢serÚa posible estimar cuÃntas especies no habÚan sido recogidas por Williams, considerando su colecciµn como una muestra aleatoria de la fauna de mariposas del archipiÕlago y conociendo la distribuciµn estadÚstica de las abundancias?. Aunque la matemÃtica asociada al problema es relativamente complicada, el enfoque de Fisher para resolverlo, fue realmente sencillo. Fisher construyµ un diagrama en el que representµ el nºmero de individuos, frente a la cantidad de especies representadas en cada clase de abundancia. La figura 3 muestra un ejemplo de distribuciµn de abundancia que sigue el modelo de la serie logarÚtmica.
|
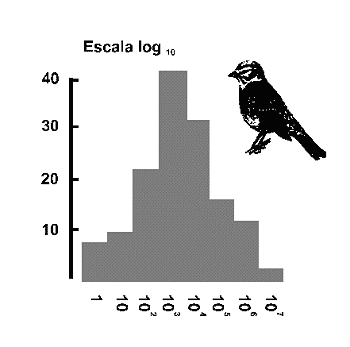
Figura 4. Distribuciµn Log normal. Las clases de abundancia (eje 'X') se representan en incrementos de orden de magnitud 1, 10, 100. 1000, 10000... La elecciµn de logaritmos en base 10, es mÃs apropiada para extensas serires de datos como en el ejemplo que se indica, que corresponde a la diversidad de aves en Gran Bretaþa (Williams, 1964). El eje 'Y' muestra el nºmero de especies en cada clase de abundancia. Modificado de Magurran (1988). |
Como indican Halffter & Ezcurra (1992), en el momento que Fisher, Corbet & Williams propusieron el modelo, la mayor parte de la comunidad cientÚfica interpretµ el trabajo como un divertimento acadÕmico. ¢A quiÕn le interesa estimar el nºmero de especies que no conoce?. La perspectiva parecÚa casi metafÚsica. La relevancia del modelo de las series logarÚtmicas, reside en que es capaz de predecir la cantidad de especies en una muestra, conociendo la cantidad de individuos que tiene cada especie en la muestra y sumando los individuos de todas las especies, como una funciµn del tamaþo total de la muestra.
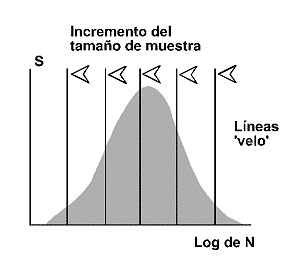
El desarrollo de Fisher y colaboradores, pronto llamµ la atenciµn de un ingeniero inglÕs radicado en Pensilvania, Frank Preston. El modelo de Preston (1948) o modelo lognormal predice que la cantidad de especies presentes en una comunidad tendrà una relaciµn Gausiana o Normal con el logaritmo de la abundancia1 (Figura 4). La mayorÚa de las comunidades estudiadas por los ecµlogos, muestran un patrµn de distribuciµn de especies-abundancia del tipo log normal. Como es bien sabido, la distribuciµn log normal es simÕtrica, en forma de campana. Asimismo, estimas repetidas de los Úndices de diversidad muestran una distribuciµn lognormal (Magurran, 1988). Sin embargo, si los datos como es frecuente, derivan de una muestra finita, la parte izquierda de la curva que representan las especies mÃs raras de la comunidad (las que no se recogen habitualmente en las muestras), està oculta. Es decir, los datos reales se ajustan, por lo general, a una distribuciµn log normal truncada (Figura 5). Cuanto mÃs pequeþa es la muestra, mÃs desplazada hacia la derecha de la ‘moda’ se sitºa la que Preston denominµ ‘veil-line’. Sµlo conocemos la porciµn de la curva a la derecha de esta lÚnea, el resto de la distribuciµn a la izquierda de dicha lÚnea, queda oculta por la ‘veil line’ (Figura 6). A medida que se incrementa el tamaþo de la muestra, la ‘lÚnea velo’ se desplaza hacia la izquierda mostrando, en primer lugar, el valor modal de la curva y, eventualmente, la distribuciµn log normal completa. Esto ºnicamente sucede en colecciones de datos que cubren un amplio rango geogrÃfico.
|
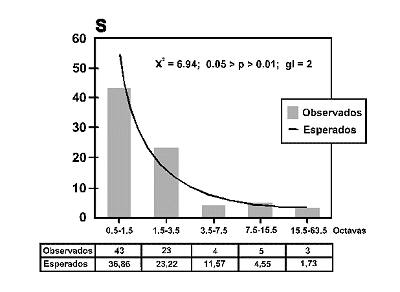
Figura 5. Distribuciµn de frecuencias por clases de abundancias u Octavas en una comunidad coleopterolµgica tropical (SerranÚa de Chiribiquete; CaquetÃ, Colombia), S: Nºmero de especies en una octava. La tabla muestra los valores observados y esperados segºn predice la distribuciµn log normal (truncada). Datos de MartÚn Piera y FernÃndez Torres (1996). |
Figura 6. Distribuciµn log normal. en muestras pequeþas ºnicamente es visible la porciµn de la distribuciµn a la derecha de la moda. A medida que se incrementa el tamaþo de la muestra, la lÚnea 'velo' se mueve de izquierda a derecha descubriendo la moda y, eventualmente, la distribuciµn log normal completa. S: nºmero de especies. Log de N: logaritmo de la abundancia. Modificado de Magurran (1988). |
Una de las consecuencias mÃs importantes del modelo log normal de Preston es que al igual que el modelo de la log-serie de Fisher, es capaz de describir la cantidad de especies que se encuentran en un Ãrea, conociendo la cantidad de individuos en el total de la muestra. La diferencia reside en que el modelo de Fisher predice un rango muy amplio de curvas especies-Ãrea, en cambio el modelo de Preston predice una relaciµn del tipo:s = k Az donde ‘k’ y ‘z’ son coeficientes derivados del modelo; ‘k’ es una constante que depende del tipo de organismos y ‘z’ es la pendiente de la recta que toma un valor muy cercano a 0,23 en muestras relativamente grandes y prµximo a 0,25 en muestras pequeþas; ‘s’ es el nºmero de especies y ‘A’ el tamaþo del Ãrea.
|
Figura 7. Relaciµn especies-Ãrea en comunidades ibÕricas de Escarabeidos coprµfagos (Coleoptera): Scarabaeidae, Aphodiinae y Geotrupinae. Nµtese que el valor del exponente 'z' se sitºa muy por debajo del valor habitual (0.23): z=0.07822. CortesÚa de Jorge M. Lobo. Datos de 25 estudios ibÕricos. |
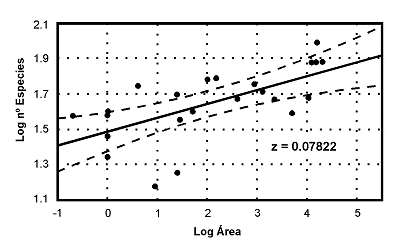
Muchos investigadores se dedicaron a poner a prueba las ideas de Preston, a travÕs del sencillo procedimiento de representar en escala logarÚtmica el Ãrea de una muestra con relaciµn al nºmero de especies contenidas en ella. Se ha encontrado que, con asombrosa regularidad, aparecÚan rectas con pendientes cercanas al valor predicho por Preston (z = 0,23) y pendientes algo mayores en islas muy depauperadas o en comunidades muy empobrecidas en especies. No obstante en algunos grupos de coleµpteros de biologÚa muy particular, como los escarabeidos coprµfagos, los valores del exponente ‘z’ encontrados son muy bajos. En muy pocas ocasiones, se han obtenido los que predice el modelo de Preston (Figura 7). La interpretaciµn mÃs plausible es que las comunidades de Scarabaeidae coprµfagos en la PenÚnsula IbÕrica, se encuentran saturadas de especies y en ellas, apenas existe el carÃcter de ‘insularidad’. Se trata ademÃs de un grupo de organismos, en el que, frecuentemente, se encuentra un patrµn de distribuciµn de especies-abundancia que no se ajusta a una distribuciµn log normal sino bimodal.
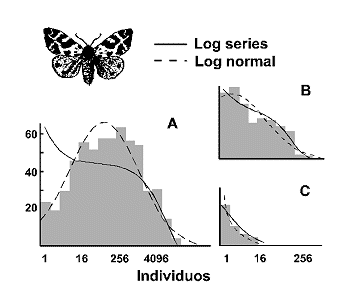
Antes de terminar con el tema de los modelos de distribuciµn de especies-abundancia, ha de comentarse que muchos datos pueden describirse igualmente con ambos modelos: log series y log normal y, en ocasiones, resulta difÚcil decidir cual de los dos se ajusta mejor a nuestros datos. La figura 8 muestra que cuando los datos se ajustan a una distribuciµn log normal truncada, es casi indistinguible de la serie logarÚtmica.
|
Figura 8. Log series y distribuciµn log normal refereidas a datos de diversidad en mariposas nocturnas. A: Abundancia de mariposas en 255 localidades de Gran Bretaþa. B: Muestra anual tÚpica en una localidad rural. C Muestra de una localidad urbana muy empobrecida. Modificado de Magurran (1988); datos de Taylor (1978). |
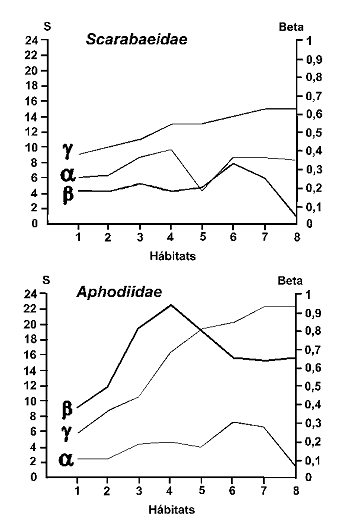
Figura 10. Variaciµn de la alfa, beta y gamma diversidad en comunidades de Scarabaeoidea coprµfagos del Parque Nacional de Doþana. S: Nºmero de especies. Beta: ëndice de diversidad Ô de Whittaker (1960). Gamma diversidad representada como la curva de riqueza acumulada. Datos de SanmartÚn, Lobo & MartÚn Piera (1998). |
|
Figura 9. JerarquÚa espacial de la
diversidad. Diversidad puntual: unidad espacial completamente censada una o varias veces. |
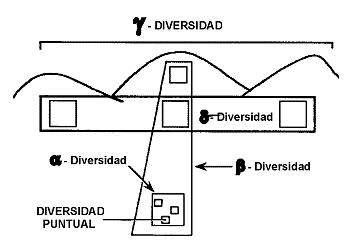
2.1.2. La JerarquÚa Espacial de la Diversidad
Tal vez la estima mÃs generalizada de
diversidad es la riqueza de especies: la popular ![]() -diversidad. Segºn Whittaker (1960 y
1975) se precisan, al menos, dos medidas adicionales para determinar la diversidad: la
diferencia entre comunidades a lo largo de un gradiente del hÃbitat
(‘Turnover’) (Ô-diversidad) y la diferencia entre Ãreas a escala
geogrÃfica (
-diversidad. Segºn Whittaker (1960 y
1975) se precisan, al menos, dos medidas adicionales para determinar la diversidad: la
diferencia entre comunidades a lo largo de un gradiente del hÃbitat
(‘Turnover’) (Ô-diversidad) y la diferencia entre Ãreas a escala
geogrÃfica (![]() -diversidad). Es la que algunos autores han denominado
jerarquÚa espacial de la diversidad (McNaughton, 1995) (Figura 9).
-diversidad). Es la que algunos autores han denominado
jerarquÚa espacial de la diversidad (McNaughton, 1995) (Figura 9).
La diversidad-![]() , es una
funciµn de la cantidad de especies presentes en un mismo hÃbitat y es el componente de
la Biodiversidad mÃs importante y mÃs comºnmente citado en las selvas tropicales
hºmedas y en los arrecifes coralinos. La Ô y
, es una
funciµn de la cantidad de especies presentes en un mismo hÃbitat y es el componente de
la Biodiversidad mÃs importante y mÃs comºnmente citado en las selvas tropicales
hºmedas y en los arrecifes coralinos. La Ô y ![]() -diversidad son tan
importantes como la riqueza a la hora de determinar la diversidad local y regional. La Ô-diversidad
es una medida del relevo faunÚstico (‘turnover’) a lo largo de un gradiente:
entre muestras de un mismo hÃbitat, entre hÃbitats o entre hÃbitats a medida que se
amplÚa el rango geogrÃfico (
-diversidad son tan
importantes como la riqueza a la hora de determinar la diversidad local y regional. La Ô-diversidad
es una medida del relevo faunÚstico (‘turnover’) a lo largo de un gradiente:
entre muestras de un mismo hÃbitat, entre hÃbitats o entre hÃbitats a medida que se
amplÚa el rango geogrÃfico (![]() -diversidad). Cuanto menor es el nºmero de especies compartidas entre dos
muestras, hÃbitats, regiones, es decir, cuanto mayor es el relevo faunÚstico
(complementariedad), mayor es la Ô-diversidad. La
-diversidad). Cuanto menor es el nºmero de especies compartidas entre dos
muestras, hÃbitats, regiones, es decir, cuanto mayor es el relevo faunÚstico
(complementariedad), mayor es la Ô-diversidad. La ![]() -diversidad es la riqueza
de especies a lo largo de un rango de hÃbitats (diversidad regional) y puede medirse como
la curva de riqueza acumulada (Figura 10), o bien, como el
producto:
-diversidad es la riqueza
de especies a lo largo de un rango de hÃbitats (diversidad regional) y puede medirse como
la curva de riqueza acumulada (Figura 10), o bien, como el
producto: ![]() =
= ![]() x Ô x
Nºmero de hÃbitats (ecosistemas; unidades paisajÚsticas, etc.).
x Ô x
Nºmero de hÃbitats (ecosistemas; unidades paisajÚsticas, etc.).
|
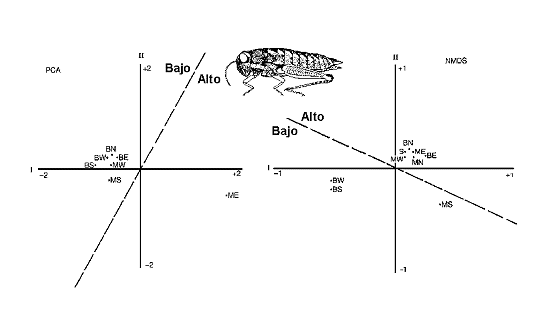
Figura 11. AnÃlisis de comunidades de ortµpteros en una reserva sudafricana, aplicando dos mÕtodos de ordenaciµn: AnÃlisis de Componentes Principales (PCA) y un mÕtodo no paramÕtrico (NMDS: ‘non metric multidimensional scaling’). PCA agrupa seis estaciones con densidades bajas de ortµpteros en tanto que NMDS, agrupa seis estaciones con altas densidades. Las siglas representan estaciones de muestreo. Modificado de Samways (1994). Datos de Samways (1990). |
La expresiµn original de Whittaker
(1960) es ÔW = S/![]() - 1, donde ‘S’ es el
nºmero total de especies en la comunidad y ‘
- 1, donde ‘S’ es el
nºmero total de especies en la comunidad y ‘![]() ’
es el promedio de especies en todos los hÃbitats considerados. Realizando este mismo
cÃlculo entre pares de hÃbitats contiguos, obtenemos la medida del reemplazamiento
faunÚstico a lo largo de todo el transecto analizado.
’
es el promedio de especies en todos los hÃbitats considerados. Realizando este mismo
cÃlculo entre pares de hÃbitats contiguos, obtenemos la medida del reemplazamiento
faunÚstico a lo largo de todo el transecto analizado.
Cody (1975) se interesµ mÃs por el cambio en la composiciµn de la comunidad, a lo largo de un gradiente del hÃbitat. Por ello su expresiµn toma en cuenta el nºmero de especies singulares, no compartidas, entre cada hÃbitat. La expresiµn de Cody es:
ÔC = g(H) + l(H)/2
siendo g(H) el nºmero de especies ganadas y l(H) el nºmero de especies perdidas a lo largo del transecto.
2.2. Medidas Evolutivas de Diversidad
Como acabamos de ver, la literatura està plagada de Úndices que miden la diversidad de las comunidades. Unos han tenido mayor aceptaciµn que otros, pero todos han tratado de expresar en un ºnico valor numÕrico, la riqueza y la dominancia/equitatividad de las comunidades en estudio. No es que el intento haya sido vano, pero tal vez sÚ algo pretencioso y demasiado simplista.
Se ha querido encontrar un Úndice que exprese la complejidad de los ecosistemas con una sola cifra y permita, ademÃs, hacer comparaciones entre diferentes lugares y/o entre distintas comunidades. Desafortunadamente no pasan de ser meros nºmeros, con poco o nulo valor conceptual. De hecho, en algunos casos se puede obtener el mismo valor de diversidad con diferentes combinaciones de valores de riqueza, abundancia y equitatividad (e.g. C2, C4 y C5 en la Tabla1) y lo que es aºn peor, una comunidad con gran riqueza de especies pero desigual distribuciµn de abundancia, tendrà valores de diversidad menores que otra pobre en especies, pero con una alta equitatividad (e.g. C1 y C3 en la Tabla 1).
Tabla 1
Variaciµn del Úndice de Diversidad H’
(Shannon-Wienner) en comunidades hipotÕticas (C1.... C5). N:
Abundancia. Sp1.... Sp11 especies.
| Sp1 | Sp2 | Sp3 | Sp4 | Sp5 | Sp6 | Sp7 | Sp8 | Sp9 | Sp10 | Sp11 | N | H’ | |
| C1 | 2 | 2 | 2 | 6 | 1,099 | ||||||||
| C2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 18 | 2,197 | ||
| C3 | 90 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 100 | 0,555 |
| C4 | 20 | 20 | 8 | 8 | 8 | 8 | 8 | 5 | 5 | 5 | 5 | 100 | 2,250 |
| C5 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 5 | 5 | 100 | 2,372 |
Por otro lado, diferentes Úndices proporcionan diferentes resultados, utilizando los mismos datos. Incluso las tÕcnicas mÃs complejas de anÃlisis multivariante (PCA - AnÃlisis de Componentes Principales -, NMDS - ‘Non-metric multiple dimensional scaling’ -, AnÃlisis de Correspondencia, AnÃlisis Canµnico, ‘Twispan’, etc.) pueden dar resultados que sugieren interpretaciones diferentes. En el ejemplo que muestra la figura 11, PCA agrupa los sitios con menores densidades de ortµpteros en tanto que NMDS, el otro mÕtodo, agrupa los de mayor densidad.
Las medidas de diversidad de cualquier tipo, tampoco nos dicen nada acerca de las relaciones funcionales de los organismos en el seno de los ecosistemas ni de la respuesta a las condiciones cambiantes del medio. El punto crucial del actual problema de perdida global de diversidad, no es sµlo quÕ hay, dµnde se encuentra, cµmo se estructura, sino tambiÕn quÕ està pasando a tenor de los cambios a pequeþa y gran escala en la superficie y en la atmµsfera del planeta.
Hay dos extremos de equivalencia funcional: 1) Cada especie en el ecosistema juega un papel relativo importante, de tal manera que la desapariciµn de cada especie debilita la integridad del ecosistema (‘Hipµtesis del Remache’). El sÚmil que plantea esta hipµtesis supone que cada especie representa algo similar a los ‘remaches’ de los antiguos aeroplanos. A medida que ‘el viejo aeroplano pierde remaches’ aumenta el peligro de que su estructura se debilite hasta no poder volar. En tÕrminos ecolµgicos esto equivale a decir que cada especie, ocupa un nicho ecolµgico ºnico e irremplazable. 2) El ecosistema està compuesto de grupos funcionales parcialmente redundantes, cada uno con especies ecolµgicamente equivalentes. De esta forma, podrÚamos perder algunas especies sin que se resienta el ecosistema en su conjunto (Hipµtesis de las especies redundantes).
AsÚ por ejemplo, en las comunidades de escarabeidos coprµfagos, grupo de coleµpteros muy diversificado, especialmente en el continente africano, se han descrito hasta siete grupos funcionales (Doube, 1991; figura 12). Es muy probable que la pÕrdida por extinciµn, migraciµn, o cualquier otra causa, no afecte sensiblemente a la estructura y funcionalidad de la comunidad, asÚ como a su participaciµn en los flujos de materia y energÚa que tiene lugar en el microecosistema excremento; pero en todo caso, habrÚa de demostrarse.
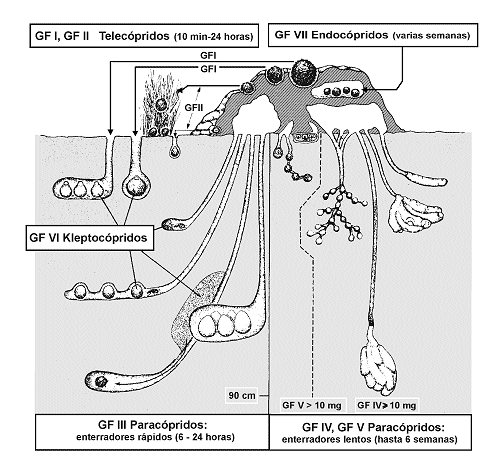 Fig. 12. Representaciµn esquemÃtica de grupos funcionales (GF) en Scarabaeoidea (Col.) coprµfagos. Modificado de Doube (1990). |
Como han seþalado Chapin et al., (1992), probablemente la realidad se encuentra en algºn punto intermedio de estos extremos. Precisamente uno de los retos de la Conservaciµn es predecir si existen especies que constituyen autÕnticas ‘piedras angulares’ del ecosistema. Es decir aquellas especies claves, no redundantes desde el punto de vista funcional que juegan un papel esencial en la integridad del sistema.
En suma, la distribuciµn de la riqueza en categorÚas de abundancia, los Úndices de diversidad, los modelos de dominancia-abundancia, los anÃlisis multivariantes son todos ellos mÕtodos descriptivos, pero ninguno explica los patrones de diversidad y sus causas. Y lo que es aºn mÃs importante, tratan a todas las especies de igual manera; no hay diferencias taxonµmicas (evolutivas) ni funcionales. Actualmente, sin embargo, tanto ecµlogos como conservacionistas coinciden en la necesidad de tomar en consideraciµn los patrones y procesos histµricos que determinan el grado de diferencia taxonµmica entre los organismos a la hora de explicar las causas que determinan la diversidad (IUCN, 1990; Cornell & Lawton, 1992; Grombridge, 1992; Haydon et al., 1993; Pearson & Juliano, 1993; Farrell & Mitter, 1993; Brooks & McLennan, 1991 y 1993; Cadle & Greene, 1993; Valkenburg & Janis, 1993; Ricklefs & Schluter, 1993; Lobo, 1998).
Tres nuevas aproximaciones se han sugerido a lo largo de esta dÕcada para cuantificar la singularidad taxonµmica (‘distinctiveness’) o grado de independencia en la historia evolutiva de un taxon particular (IEH: ‘independent-evolutive-history’) (May, 1996).
i ) Medidas Moleculares (Embley et al., 1996).
ii) Medidas expresadas en tÕrminos de Riqueza Taxonµmica
iii) Medidas FilogenÕticas.
A continuaciµn me referirÕ a las dos aproximaciones con las que estoy mÃs familiarizado; las dos ºltimas, especialmente la tercera. Antes quisiera hacer una breve disgresiµn conceptual que conviene tener siempre presente cuando trabajamos con estimaciones fundamentadas en una visiµn taxonµmica de la diversidad: me refiero al concepto de especie.
Los mºltiples conceptos de especie en diferentes grupos e incluso dentro del mismo grupo de organismos, son una causa importante de incertidumbre inherente a todos los aspectos de la investigaciµn sobre biodiversidad que utilizan la especie como ‘unidad estÃndard’. Si la unidad de medida es variable, las conclusiones basadas en ella han de tomarse con precauciµn. El concepto biolµgico de especie se ha utilizado como una entidad comparable en diferentes organismos, pero esta comparaciµn no siempre es posible entre organismos evolutivamente muy dispares. ¢Cµmo se compara, por ejemplo, un insecto y un hongo cuando se examinan con mÕtodos moleculares o filogenÕticos? ¢cuÃl es la fiabilidad de las mºltiples especies definidas ºnicamente mediante ejemplares preservados en colecciones?, ¢cµmo aplicar el concepto biolµgico de especie a organismos que nunca han experimentado reproducciµn sexual?. Conviene meditar y tener siempre presente estas cuestiones.
2.2.1. Diversidad Taxonµmica
Como ya he indicado, la estima mÃs
generalizada de diversidad es la riqueza de especies, la popular ![]() -diversidad.
Ahora bien, a medida que se incrementa el nºmero de especies, cabe esperar que,
paralelamente, se incremente la presencia de grupos taxonµmicos divergentes de alto
rango. Se discute si el nºmero de taxones de alto rango, pueden proporcionar una medida
de diversidad mÃs apropiada que el nºmero de especies. En efecto, desde una perspectiva
estrictamente conservacionista, cabe preguntarse si el nºmero de Clases, Ordenes,
Familias, ..., presentes en un Ãrea, es una estima conveniente y rÃpida del nºmero de
especies. La pregunta tiene profundas implicaciones prÃcticas si tenemos en cuenta la
ingente cantidad de trabajo taxonµmico descriptivo que aºn queda por hacer en extensas
regiones del Planeta, especialmente en las Ãreas tropicales. En efecto, si esta relaciµn
fuera de carÃcter universal, podrÚamos ‘subrogarnos’ a estos taxones
superiores, para obtener estimas prÃcticas y rÃpidas de diversidad, sin esperar a
concluir el Inventario exhaustivo de especies. No obstante, los datos no son concluyentes.
-diversidad.
Ahora bien, a medida que se incrementa el nºmero de especies, cabe esperar que,
paralelamente, se incremente la presencia de grupos taxonµmicos divergentes de alto
rango. Se discute si el nºmero de taxones de alto rango, pueden proporcionar una medida
de diversidad mÃs apropiada que el nºmero de especies. En efecto, desde una perspectiva
estrictamente conservacionista, cabe preguntarse si el nºmero de Clases, Ordenes,
Familias, ..., presentes en un Ãrea, es una estima conveniente y rÃpida del nºmero de
especies. La pregunta tiene profundas implicaciones prÃcticas si tenemos en cuenta la
ingente cantidad de trabajo taxonµmico descriptivo que aºn queda por hacer en extensas
regiones del Planeta, especialmente en las Ãreas tropicales. En efecto, si esta relaciµn
fuera de carÃcter universal, podrÚamos ‘subrogarnos’ a estos taxones
superiores, para obtener estimas prÃcticas y rÃpidas de diversidad, sin esperar a
concluir el Inventario exhaustivo de especies. No obstante, los datos no son concluyentes.
Algunos autores argumentan que sigue siendo necesario dirigir nuestra atenciµn a las especies si pretendemos evaluar y planificar la conservaciµn de la biodiversidad. AsÚ, Prance (1996) constata que de las 292 familias de plantas neotropicales, ºnicamente 40 son exclusivas o casi exclusivas de esa regiµn. Estas 40 familias reºnen tan sµlo 5,777 especies, es decir, el 6.4% de las especies neotropicales conocidas. Por el contrario, sµlo dos familias con una radiaciµn explosiva, BromeliÃceas y CactÃceas, reºnen 4,000 especies. Asimismo, analizando la relaciµn especies/gÕneros en algunas familias pantropicales (ChrysobalanÃceas), en los inventarios florÚsticos de distintos continentes (êfrica y SudamÕrica) y los catÃlogos de algunas regiones tropicales bien inventariadas tales como Perº, Prance termina concluyendo que la riqueza especÚfica es el mejor indicador de la diversidad total de una regiµn
Otros autores, sin embargo, encuentran evidencias de una alta correlaciµn en diferentes regiones, entre el nºmero de familias y el nºmero de especies en diferentes grupos de organismos (Figura 13): helechos, plantas con semillas terrestres y acuÃticas, insectos, anfibios, reptiles y mamÚferos (Williams & Gaston, 1994; Williams et al., 1994 a, 1997 a). De esta forma, sugieren que la riqueza de taxones de alto rango, puede representar una estima adecuada de la diversidad que, en ºltima instancia, representa la riqueza de especies e incluso mÃs remotamente, la riqueza genotÚpica (genes) y fenotÚpica (caracteres).
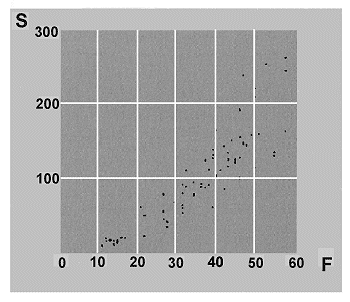
2.2.2. Medidas de diversidad filogenÕtica
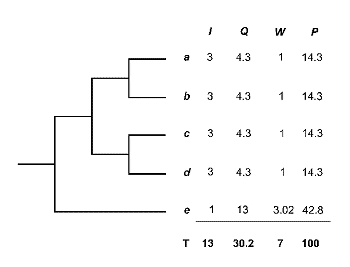
En el transcurso de esta dÕcada, sistemÃticos y biogeµgrafos vienen proponiendo nuevas medidas de diversidad que cuantifican la diferencia o singularidad (‘distinctiveness’) taxonµmica de los organismos. Estas medidas se fundamentan en las relaciones genealµgicas (ancestro-descendiente) entre organismos, en relaciµn con diferentes modelos de evoluciµn de genes o caracteres (Vane-Wright et al., 1991; Williams et al., 1994 b; Williams & Humphries, 1994; Humphries et al., 1995). El mÕtodo inicial de Vane-Wright, Humphries & Williams (1991) modifica parcialmente la propuesta de Mickevich y Platnick (1989). Segºn estos autores, el contenido de informaciµn de una clasificaciµn jerÃrquica està determinado por el nºmero de nodos del cladograma. Es decir, incluso con igual nºmero de taxones, la cantidad de informaciµn varÚa en funciµn de la topologÚa del cladograma.
El procedimiento de estos autores consiste en asignar un valor objetivo a la singularidad taxonµmica o grado de independencia en la historia evolutiva de un taxon. El mÕtodo de Vane-Wright, Humphries and Williams (1991) es simple. El cladograma de la figura 14 lleva asociadas cuatro columnas. La primera (I) indica el contenido de informaciµn filogenÕtica de cada taxon (a en 3 grupos; b en otros tres; c: 3; d: 3 y e: 1; Total=13). La segunda columna (Q) contiene los cocientes del total de informaciµn (13) entre cada parcial (13:3=4.3; 13:3; 13:3; 13:3 y 13:1=13). En la tercera columna (W) se ha estandarizado dividiendo por el menor valor (4.3). La cuarta columna (P) expresa el porcentaje de Informaciµn. Como se puede apreciar, el taxon ‘e’, es el que arroja mayor valor. AsÚ pues, cuanto menor es el nºmero de nodos, es decir el nºmero de cladogÕnesis y, por tanto, mÃs primitivo es el taxon en una filogenia, mayor es su porcentaje de Informaciµn, Peso TÃxico o Singularidad FilogenÕtica, ya que la proporciµn en la que contribuye a la diversidad total del cladograma (contenido de informaciµn) es mayor.
Esta medida de singularidad filogenÕtica o Peso TÃxico, como la denominaron Vane-Wright, Humphries & Williams (1991), cumple dos condiciones esenciales: i) La equivalencia taxonµmica (igual rango) viene expresada por idÕntico valor y ii) La singularidad taxonµmica varÚa gradualmente a lo largo del cladograma: no todas las especies son iguales. Sin embargo Williams et al. (1993) han argumentado que, en contra de las expectativas iniciales, la maximizaciµn de la divergencia filogenÕtica (primitiva) precoz (‘root weight’), no satisface los valores intuitivos de todos los conservacionistas. Por ello, a lo largo de estos ºltimos aþos, han surgido nuevos refinamientos metodolµgicos que cuantifican la singularidad taxonµmica y la diversidad filogenÕtica de los organismos, a travÕs de los rasgos o caracteres - bioquÚmicos, estructurales, ecolµgicos, comportamentales,..., - que expresan la variabilidad genÕtica de los organismos y el patrµn de divergencia evolutiva representado en los cladogramas. Es decir, los caracteres fenotÚpicos (y/o los genes que los codifican) proporcionan una ‘divisa’ del valor de diversidad.
Una ventaja de esta aproximaciµn es que proporciona una visiµn unificada de los tres niveles tradicionales de diversidad, por cuanto utiliza una expresiµn de la diversidad genÕtica (los caracteres) como base para valorar tanto la diversidad de especies, por su riqueza relativa en genes diferentes, como la diversidad de ecosistemas, por la riqueza relativa de procesos a los que, en ºltimo tÕrmino, contribuyen los propios genes.
En resumen, puesto que los genes y los caracteres que codifican son heredables, los biµlogos podemos estimar la biodiversidad mediante medidas taxonµmicas o filogenÕticas, utilizando nuestro conocimiento de las relaciones genealµgicas (jerÃrquicas) entre organismos en combinaciµn con modelos (anagenÕticos, cladogenÕticos) de evoluciµn de genes y caracteres (Williams et al., 1994 b; Humphries et al., 1995).
Entre estas medidas vamos a fijarnos esencialmente en dos: la ‘Riqueza de Caracteres’ y la ‘Combinaciµn de Caracteres’ y, mÃs brevemente, en otras dos derivadas de ellas: la Diversidad FilogenÕtica (PD) y la ‘Longitud del subÃrbol expandido’. Otras mÃs refinadas y tambiÕn algo mÃs complejas tales como la ‘diversidad p-mediana’ y la ‘diversidad p-mediana discreta’, exceden los lÚmites de este artÚculo. El lector interesado en profundizar en el tema puede consultar los trabajos de Faith (1992 a y b; 1994 y 1996); Humphries et al., (1995); Williams (1993); Williams & Humphries (1994 y 1996) y Williams et al., (1994 b) y bibliografÚa en ellos citada.
2.2.2.1. Riqueza de Caracteres
Cuando se trabaja con un gran nºmero de
especies, el valor de la riqueza de especies (![]() -diversidad), tiende a ser una expresiµn razonable de la
variabilidad genÕtica y de la riqueza de caracteres en que aquÕlla se expresa. Esta
relaciµn ha sido ilustrada en varios organismos: insectos, plantas neotropicales y aves
(Williams & Humphries, 1996) (Figura 15).
-diversidad), tiende a ser una expresiµn razonable de la
variabilidad genÕtica y de la riqueza de caracteres en que aquÕlla se expresa. Esta
relaciµn ha sido ilustrada en varios organismos: insectos, plantas neotropicales y aves
(Williams & Humphries, 1996) (Figura 15).
|
Figura 15. Algunos ejemplos de relaciµn entre el nºmero de especies (S) y la riqueza de caracteres (C). a: 241 especies mundiales de abejorros del gÕnero Bombus (Apidae); coeficiente de correlaciµn de Spearman=0.986; datos de distribuciµn y clasificaciµn de Williams (1991 y 1995). b: 729 especies neotropicales de las familias Dichapeta-lÃceas, LecythidÃceas, CaryocarÃceas, ChrysobalanÃceas y el gÕnero Panopsis; coeficiente de correlaciµn de Spearman=0.994; datos de Williams/Prance et al. (en prensa). c: 218 especies de aves nidificantes en Gran Bretaþa; coeficiente de correlaciµn de Spearman=0.952; datos Gibbons et al., 1993; Williams/Gibbons et al. (en prensa). Modificado de Williams & Humphries (1996). |
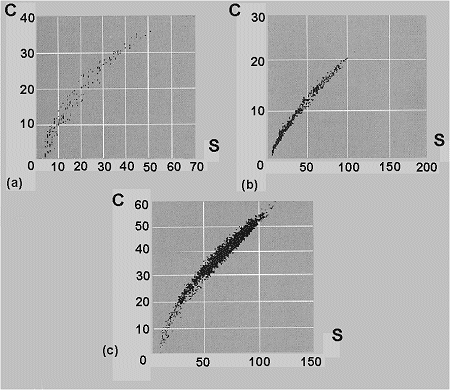
Por consiguiente, utilizando clasificaciones, preferiblemente ilustradas como cladogramas, podemos obtener una estimaciµn de la diversidad a travÕs de la riqueza de rasgos-atributos-caracteres que identifican a los organismos. El modelo asume que todos los rasgos se originan y persisten en todos los descendientes del linaje y ademÃs, que el patrµn de ramificaciµn cladogenÕtica y la longitud de la(s) rama(s) expresada como el nºmero de rasgos que representan novedades evolutivas en un linaje, proporciona una estima adecuada de diversidad (Faith, 1996).
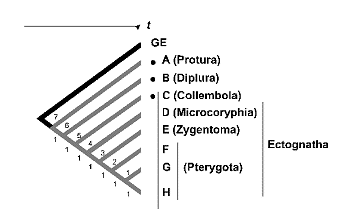
La estimaciµn de la diversidad mediante la riqueza de caracteres responde a un modelo anagenÕtico de evoluciµn, en el que se acepta una relaciµn directa entre la longitud de las ramas que configuran los diferentes linajes y la antig■edad relativa de los taxones. Este es el modelo propuesto comºnmente en la evoluciµn de los atributos moleculares, conocido como ‘reloj molecular’. Si aceptamos esta idea, todos los taxones podrÚan situarse sobre el Ãrbol filogenÕtico a igual distancia de la raÚz, convirtiendo el cladograma en un Ãrbol ultramÕtrico. Esta equidistancia, se refiere no sµlo al eje del tiempo sino tambiÕn al nºmero de cambios o novedades evolutivas (caracteres) representadas sobre la rama. De la figura 16 se desprende que en Ãrboles ultramÕtricos, la diversidad medida como la riqueza de caracteres dentro de un conjunto de taxones, es mÃxima en aquellos taxones de divergencia filogenÕtica precoz o mÃs antigua. Desde esta perspectiva, bastarÚa conservar A (e.g.: una especie de Proturo), B (e.g.: Dipluro) y C (e.g.: ColÕmbolo) a pesar de que los taxones D -- H pudieran representar los restantes Ectognatos (D, e.g.: Microcoryphia; E, e.g.: Zygentoma y de F a H e.g.: diferentes grupos de Pterygotas). El mismo sesgo del que adolecÚa el peso tÃxico, la medida original de Vane-Wright et al. (1991).
Una aproximaciµn mÃs consistente fundamentada tambiÕn en la longitud de las ramas que configuran el cladograma, es la formulaciµn original de Diversidad FilogenÕtica (Faith, 1992 a y b, 1994 y 1996). La diversidad filogenÕtica (PD) de un subgrupo de taxones se define como la suma de las longitudes de las ramas que a lo largo del Ãrbol, conectan todos los taxones de un subgrupo de organismos, en dicho Ãrbol, a travÕs del ‘camino mÃs corto’ (‘minimum spannign path’). Desde esta perspectiva, el patrµn filogenÕtico para el grupo taxonµmico en cuestiµn, nos da una predicciµn de los patrones subyacentes de diversidad de rasgos. En efecto, un subgrupo de taxones que evidencian una mayor proporciµn del Ãrbol, es, en este sentido, mÃs diverso (Figura 17).
Esta definiciµn de la diversidad filogenÕtica origina otro concepto esencial en la cuantificaciµn de la diversidad de los organismos: El concepto de Complementariedad o Incremento de la Diversidad FilogenÕtica, I(x) (Faith, 1992 a; 1994, 1996). La complementariedad en este contexto, se refiere a nuevos rasgos o atributos con los que un grupo adicional de especies puede contribuir a la diversidad del Ãrbol filogenÕtico. Este reemplazamiento (‘turnover’) en rasgos, denominado por algunos autores Q-diversidad (Faith, 1996), juega asÚ el mismo papel que la Ô-diversidad en un contexto ecolµgico.
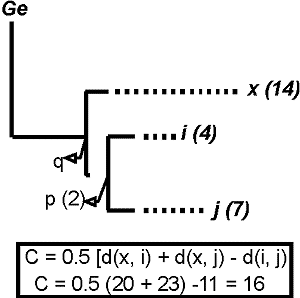
La medida de complementariedad puede ser fÃcilmente calculada utilizando las distancias entre pares de especies (Faith, 1992 a y 1994). Por ejemplo, para el Ãrbol hipotÕtico de la figura 18 el incremento de la diversidad o complementariedad cuando se aþade la especie ‘x’ al subgrupo ‘i’ ‘j’, viene dado para esta sencilla expresiµn:
C= 0.5 [( d (x, i) + d (x, j) - d (i, j)] C= 0.5 [( d (x, i) + d (x, j) - d (i, j)] (1) (Faith, 1996)
donde d(x, i) y d(x, j) son las distancias entre los pares de especies medidas como nºmero de rasgos a lo largo de las ramas y d(i,j), los rasgos distintos en ‘i’ y ‘j’. En realidad, ‘C’ mide la distancia desde la especie ‘x’ a la porciµn del Ãrbol representado por el par ‘i’ ‘j’, es decir, la distancia entre ‘x’ y el ancestro ‘p’. Esta distancia representa el incremento en diversidad filogenÕtica, I(x), resultante de la adiciµn de un nuevo taxon:
I (x) = MÚnimo sobre todos los i, j
en ‘s’ 0.5 (Dx,i + Dx,j - Di,j) (2) (Faith, 1994).
siendo Dx,i, Dx,j y Di,j,, como acabamos de ver, las distancias entre los pares de especies. Esta fµrmula, en realidad, mide la longitud extra del Ãrbol al aþadir la rama del taxon ‘x’ al subgrupo ‘s’ de taxones ‘i’ ‘j’.
Si no asumimos una relaciµn directa entre el nºmero de cambios o novedades evolutivas (caracteres, atributos) y el tiempo evolutivo (antig■edad versus modernidad de los taxones), la riqueza de caracteres y, por tanto, la longitud de las ramas, podrÚa no ser una estima adecuada de diversidad. Ahora bien, podemos aceptar que, en cambio, existe una correlaciµn entre el nºmero de cambios y el nºmero de nodos en el cladograma. En tal caso, el patrµn de ramificaciµn y por tanto, el valor predictivo de los nodos (especialmente para la diversidad morfolµgica), podrÚa ser aºn mÃs elevado que la longitud de las ramas de los linajes (Williams et al., 1994 b).
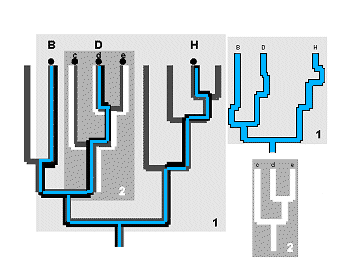
La medida mÃs evidente de representatividad taxonµmica, es la longitud del subÃrbol expandido (‘spanning subtree length’ o ‘cladistic path length’) (Williams et al., 1993). La longitud del subÃrbol expandido, expresa simplemente la fracciµn de la clasificaciµn que està representada en el cladograma de un subgrupo de taxones (Williams & Humphries, 1994) (Figura 19) y mide la divergencia taxonµmica entre taxones de una biota sumando el nºmero de nodos de la clasificaciµn que median entre todos los nodos incluidos en la biota, contando cada nodo una sola vez. Se trata, por tanto, de un modelo claramente cladogenÕtico.
La longitud del subÃrbol expandido puede trabajar bien si disponemos de informaciµn suficiente sobre la longitud de las ramas del Ãrbol filogenÕtico tal como sostiene Faith (1992 a y 1994), pero sin esta informaciµn extra, la versiµn del conteo de nodos puede ser equÚvoca y no garantiza completamente la discriminaciµn de los agrupamientos taxonµmicos, tal como se le exige a cualquier medida de diversidad (Williams & Humphries, 1994).
2.2.2.2. Combinaciµn de Caracteres
Una medida alternativa a la riqueza de caracteres, consiste en evaluar la riqueza en diferentes combinaciones de caracteres. Esta medida es un refinamiento del criterio de representatividad y descansa sobre el mismo concepto de la longitud del subÃrbol expandido.
La diversidad medida a travÕs de la combinaciµn de caracteres identifica el subgrupo de organismos distribuidos mÃs regularmente sobre la topologÚa del cladograma. Este patrµn de regularidad representarà el mayor nºmero de taxones a todos los niveles dentro de la jerarquÚa taxonµmica y, por consiguiente, el mayor nºmero de combinaciones de caracteres (Humphries et al., 1995). El argumento lµgico de este criterio es el siguiente: cuanto mÃs regularmente estÃn distribuidos los taxones en el Ãrbol filogenÕtico, el subÃrbol que los contiene representa mejor la clasificaciµn general (Figura 20) y, por ello, los subÃrboles mÃs extendidos (‘dispersed’), son mÃs representativos de una variedad de subgrupos con diferentes combinaciones de caracteres.
2.2.3. Medidas FilogenÕticas e Informaciµn de las Clasificaciones
A menudo se carece de informaciµn filogenÕtica y por tanto se desconoce el patrµn de divergencia evolutiva de los organismos. En tales casos, la ºnica informaciµn disponible sobre las posibles relaciones filogenÕticas de los organismos, es la clasificaciµn taxonµmica pura y dura. Incluso en tales casos, la clasificaciµn bÃsica puede ser interpretada en un marco de diversidad filogenÕtica (Faith, 1994).
Si el promedio de divergencia taxonµmica (es decir, longitud de la rama) con relaciµn al ancestro comºn de un gÕnero es ‘dg’ y el promedio de divergencia de una especie es ‘ds’, el Incremento de Diversidad FilogenÕtica vendrà dado por:
I = G dg + N ds I = G dg + N ds (3) (Faith, 1994)
donde G es el nºmero de gÕneros previamente no representados y N el nºmero de nuevas especies presentes (Tabla 2).
Williams & Humphies (1994) han criticado esta medida por cuanto la asignaciµn de rangos taxonµmicos al margen de las relaciones de grupo hermano, es una labor decididamente arbitraria (ver tambiÕn Gauthier et al., 1988).
2.3. Medidas Corolµgicas de Diversidad: Rareza y Endemicidad
Entender el problema de la Biodiversidad, implica discutir el problema de la rareza biolµgica. En efecto, ya hemos visto que en una muestra finita, son las especies poco abundantes (raras) las que casi siempre quedan fuera del muestreo y, en la mayorÚa de los casos, la diferencia entre el nºmero de especies observadas y el nºmero real de especies que integran la comunidad, se refiere precisamente a las especies raras.
INFORMACION DISPONIBLE |
VERSION DE DIVERSIDAD FILOGENèTICA (PD) |
|
| 1 | Clasificaciµn que puede no reflejar la filogenia | I = Gdg + Nds dg, ds subjetivamente definidas; o igual peso para todas las especies. |
| 2 | Informaciµn de la Clasificaciµn concordante con la Filogenia | I = Gdg + Nds dg, ds subjetivamente definidas |
| 3 | Informaciµn de la Clasificaciµn mÃs informaciµn filogenÕtica adicional | I = Gdg + Nds dg, ds variables entre taxones de acuerdo con la informaciµn filogenÕtica. |
| 4 | Estimas filogenÕticas corroboradas. Önicamente TopologÚas. | I(x) = 0.5 (Dx,i
+ Dx,j - Di,j) Longitud de las ramas definidas por algºn modelo o iguales a alguna constante. |
| 5 | Estimas filogenÕticas corroboradas, incluyendo la longitud de las ramas. | I(x) = 0.5 (Dx,i + Dx,j - Di,j) |
Rabinowitz et al. (1986), se dedicaron a estudiar este problema, no tanto desde el punto de vista estadÚstico de Fisher y Preston, sino enfocando el problema del comportamiento ecolµgico y caracterizaciµn de las especies raras.
La Rareza de un organismo es la condiciµn de ser infrecuente y puede presentarse como rareza de ocupaciµn entre Ãreas (‘range-size rarity’) o bien como rareza de individuos dentro de un Ãrea. Sin embargo, no todas las especies con rangos geogrÃficos relativamente restringidos son localmente poco abundantes y viceversa (Williams et al., 1997 b).
Rabinowitz y colaboradores, encontraron que las causas de la rareza ecolµgica de las especies dependen, al menos, de la combinaciµn de tres factores: distribuciµn geogrÃfica, especificidad del hÃbitat y tamaþo local de la poblaciµn. De esta forma, pudieron categorizar hasta siete clases de rareza (Tabla 3). La formulaciµn conceptual de Rabinowitz y colaboradores, tiene enorme trascendencia desde el punto de vista de la BiologÚa de la Conservaciµn como veremos mÃs adelante en el caso de los Insectos. Retengamos por el momento estos tres conceptos:
Rareza BiogeogrÃfica. Se califica asÚ a las especies que colonizan un nºmero reducido de Ãreas. Aunque a menudo tienden a confundirse, rareza biogeogrÃfica y endemicidad no expresan necesariamente la misma caracterÚstica espacial. No obstante, como veremos inmediatamente, rareza biogeogrÃfica y endemismo pueden llegar a ser conceptos equivalentes.
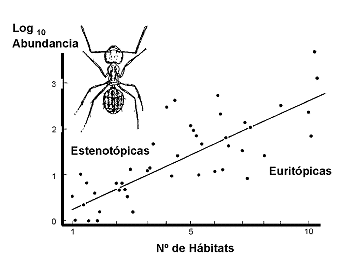
Rareza de HÃbitat. Otras especies, en cambio, son muy especÚficas en cuanto al hÃbitat, aunque muestran amplios rangos de distribuciµn geogrÃfica. Este grupo està formado por las que se conocen en EcologÚa como especies estenµicas o de hÃbitat restringido, en contraste con las euriµicas. La rareza puede estar fuertemente relacionada con la tolerancia al biµtopo. AsÚ por ejemplo, Samways (1983) encontrµ una fuerte correlaciµn positiva entre abundancia y tolerancia al biµtopo (Figura 21).
Rareza demogrÃfica. Hay especies que son demogrÃficamente raras, es decir, presentan densidades bajas en todo el Ãrea de distribuciµn, aunque Õsta sea amplia y aunque no estÕn asociadas a hÃbitats muy especÚficos.
En sentido estricto, la Endemicidad es un concepto diferente al de rareza por cuanto expresa la condiciµn de estar restringido a un Ãrea particular, de rango geogrÃfico preestablecido, dentro de un Ãrea de estudio. Sin embargo, la rareza de ocupaciµn entre Ãreas puede ser equivalente a una medida de endemicidad, ya que tambiÕn el rango geogrÃfico de la(s) especie(s) rara(s), està comprendido dentro del Ãrea de estudio y la medida del rango de rareza, es, en definitiva, una expresiµn relativa a la extensiµn total de dicho Ãrea (Williams et al., 1997 b).
En ocasiones, el calificativo endÕmico tiene connotaciones biogeogrÃfico-histµricas al referirse a elementos relictos cuyo rango geogrÃfico actual, es resultado de procesos de fragmentaciµn y/o reducciµn del Ãrea primitiva en Õpoca histµrica, mÃs o menos reciente. Pero nuevamente endemicidad y carÃcter relicto no son conceptos sinµnimos, aunque a menudo, pueden ser equivalentes. En efecto, las especies relictas no siempre estÃn constituidas por poblaciones poco abundantes, ni muestran restricciones geogrÃficas o de hÃbitats.
La endemicidad se define en un contexto geogrÃfico relacional, es decir, en comparaciµn al Ãrea de distribuciµn de otros organismos y tiene una amplia aceptaciµn entre los biogeµgrafos. En efecto, es un criterio clÃsico utilizado en el denominado AnÃlisis CrÚtico de êreas (Ackery & Vane-Wright, 1988), cuyo objetivo es la selecciµn de Ãreas de conservaciµn. Este mÕtodo trata de encontrar el mÚnimo nºmero de Ãreas que contienen, al menos, una poblaciµn de cada una de las especies analizadas, estableciendo la secuencia de Ãreas de endemicidad: primero las Ãreas con mayor nºmero de endemismos, en ºltimo lugar las que sµlo contienen uno; ver tambiÕn Vane-Wright et al (1991).
La medida de endemicidad expresada como rango de rareza espacial o rango de ocupaciµn de Ãreas, puede cuantificarse a travÕs del nºmero de parcelas ocupadas en un mapa con una proyecciµn reticulada, por ejemplo, la proyecciµn UTM; por ello, depende fuertemente de la escala espacial utilizada. AsÚ por ejemplo, Baz (1996) expresa la endemicidad como la ocupaciµn proporcional (Pj) de la especie ‘j’ en ‘n’ cuadrÚculas:
Pj = ![]() Sij/n
Sij/n
donde ‘n’ es el nºmero total de cuadrÚculas y Sij la presencia/ausencia de la especie ‘j’ en la cuadrÚcula ‘i’.
Los centros de endemismo, como las Ãreas que alojan una gran diversidad de especies u otros taxones, o las que incluyen una muestra representativa de la jerarquÚa taxonµmica y/o filogenÕtica, pueden constituir un valor de conservaciµn y se identifican como aquellas Ãreas en las que coexisten un gran nºmero de especies con distribuciones espaciales muy restringidas (Williams, 1993).
Actualmente se utilizan diferentes medidas del rango de rareza espacial para detectar las ‘Puntos CrÚticos’ o Ãreas de mÃxima endemicidad, comunes a diferentes grupos de organismos. Se usan dos aproximaciones para medir el grado de endemicidad: i) medidas discontinuas, utilizando umbrales espaciales que incluyen ºnicamente las especies mÃs raras; (e.g.: se considera endÕmico el cuartil de especies con menor rango geogrÃfico); estos umbrales se establecen arbitrariamente y ii) medidas que son funciones continuas del tamaþo del rango geogrÃfico. Estas medidas escapan a la arbitrariedad de los umbrales, pero no existe una fµrmula ‘natural’ evidente para pesar la rareza del rango (Williams et al., 1997 b).
Tabla 3
Siete clases de rareza atendiendo a tres caracterÚsticas ambientales: Distribuciµn geogrÃfica, hÃbitat y tamaþo de las poblaciones. Modificado de Rabinowitz et al.. (1986). RB: Rareza biogeogrÃfica (endemismo), RD: Rareza demogrÃfica, RH: Rareza de hÃbitat, U: Especies ubiquistas
| DISTRIBUCIÆN GEOGRêFICA: | Amplia |
Restringida | |||
| ESPECIFICIDAD DE HêBITAT | Laxa | Fuerte | Laxa | Fuerte | |
| TAMAîO DE LAS POBLACIONES LOCALES: | Grande | U | RH | RB | RB; RH |
| Pequeþo | RD | --- | RB; RD | RB; RH; RD | |
2.4. Identificando êreas de MÃxima Diversidad
La Conservaciµn es una ciencia de crisis y como tal se espera de ella decisiones, las mÃs de las veces urgentes. Desde este punto de vista es claro que necesitamos conservar y preservar rareza, riqueza, singularidad y los hÃbitats, ecosistemas y paisajes en los que estas caracterÚsticas estÕn representadas µptimamente. Pero para ello, es preciso aceptar que no todas las especies son ‘iguales’. MÃs allà de las medidas de diversidad y mÃs allà de los valores humanos imperantes en cada regiµn, precisamos reconocer y cuantificar la singularidad histµrico-evolutiva (filogenÕtica) y tambiÕn, aunque esto es bastante mÃs complejo, la funcionalidad ecolµgica de las especies.
El propµsito de la metodologÚa discutida en las secciones anteriores, es aplicar las medidas propuestas a la conservaciµn in situ de la Biodiversidad, identificando las êreas CrÚticas (‘Hot Spots’) con prioridad de acciµn, de acuerdo con el valor o combinaciµn de valores de diversidad elegidos (Vane-Wright et al., 1991; Faith, 1992 b; May 1990 a). Es mÃs razonable, se argumenta, dar mayor prioridad al Ãrea o Ãreas que alojan una representaciµn µptima de dichos valores: especies de divergencia precoz, nºmero de especies, riqueza de caracteres, combinaciµn de caracteres, endemicidad, etc.
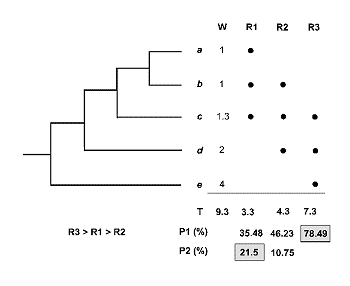
El mÕtodo inicial de Vane-Wright, Humphries and Williams (1991) identifica los ‘Puntos CrÚticos’ de diversidad, combinando la informaciµn filogenÕtica y biogeogrÃfica en cada Ãrea. En la figura 22, R1, R2 y R3 son las regiones en las que estÃn representados los taxones ‘a’, ‘b’,’ c’, ‘d’ y ‘e’. Es claro que si nos basamos en riqueza de especies ºnicamente, no podremos tomar ninguna decisiµn. De la misma forma, si tomamos en cuenta el nºmero de endemismos decidiremos que las Ãreas a conservar son las regiones R1 y R3, ambas con un endemismo; tampoco podremos priorizar ninguna de las dos. Ahora bien, si nos preguntamos cuÃl es el peso tÃxico o porcentaje de singularidad taxonµmica total en cada regiµn, entonces sÚ podremos tomar alguna decisiµn razonada. Veamos cµmo:
R1 arroja el 35% del total de informaciµn filogenÕtica (T=9.3; 1+1+1.3 = 3.3/9.3=35.48); R2 = 46% (1+1.3+2/9.3) y R3 = 78% (1.3+2+4/9.3); columna P1. Debido a la simpatrÚa de especies los totales suman mÃs del 100%. AsÚ pues, R3 es la primera elecciµn. ¢CuÃl es la segunda? De los valores anteriores cabrÚa pensar que la segunda Ãrea ha de ser R2 y la tercera R1, ya que R2 aloja mayor porcentaje de informaciµn (46,23>35,48). Ahora bien tomando en cuenta el concepto esencial de Complementariedad de faunas, R2 no es la elecciµn correcta. En efecto, si R3 mantiene un estado razonable de conservaciµn de los taxones ‘c’, ‘d’ y ‘e’, estaremos ahora interesados en conservar ‘a’ y ‘b’. Con este argumento volvemos a recalcular los porcentajes de singularidad taxonµmica y en la columna P2 anotamos que R1 (1+1/9.3 = 21.5%) en tanto que R2 (1/9.3 = 10.75%). AsÚ pues, el ranking es R3>R1>R2.
Por consiguiente, en la designaciµn de Ãreas de conservaciµn adquiere especial relevancia el concepto de Complementariedad, el cual, permite distinguir entre Ãreas irremplazables, flexibles y redundantes. Esto es importante a la hora de establecer criterios de conservaciµn por parte de los gestores del Medio Ambiente.
Como hemos visto al hablar de riqueza de rasgos/caracteres, la complementariedad es una funciµn de los atributos compartidos entre dos o mÃs conjuntos. En sentido estricto, la complementariedad se refiere a la cantidad de nuevos atributos (nºmero de especies, de caracteres, endemismos, etc.) representados en un conjunto, con los que contribuye a otro u otros conjuntos, carentes de dichos atributos.
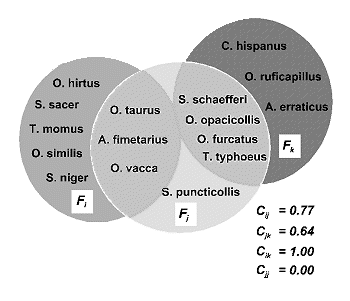
Si hablamos, por ejemplo, de especies y Ãreas, la idea se puede expresar grÃficamente tal como se ilustra en la figura 23. La complementariedad de las Ãreas que alojan a las faunas Fi y Fj es mayor que la que existe entre Fj y Fk, es mÃxima entre Fi y Fk (no comparten ninguna especie) y nula entre dos Ãreas idÕnticas. AsÚ pues, el valor de complementariedad siempre serÃ: 0 <= Cjk <= 1.
Colwell & Coddington (1994) expresan la Complementariedad (Cjk) en la ecuaciµn:
Cjk = Ujk/Sjk Cjk = Ujk/Sjk (4)
donde Ujk es el nºmero de especies ºnicas en ambas listas y Sjk la riqueza total combinada. Ujk y Sjk toman respectivamente los valores:
Ujk = Sj + Sk - 2Vjk
Sjk = Sj + Sk - Vjk
siendo Sj el nºmero de especies en la lista ‘j’; Sk el nºmero de especies en la lista ‘k’ y Vjk el nºmero de especies en comºn entre ambas listas. En la figura 23, Fi y Fk no comparten ninguna especie y, por tanto, son faunas totalmente complementarias: Sjk = 8 + 7 - 0 = 15 y Ujk = 8 + 7 - 0 = 15; asÚ pues: Cjk= Ujk/Sjk = 15/15=1. Por el contrario, si las dos listas son completamente idÕnticas (e.g.: Fj y Fj): Sjj = 8 + 8 - 8 = 8 y Ujj = 8 + 8 - 16 = 0; por tanto: Cjj= Ujj/ Sjj = 0/16=0.
Ya hemos visto que la complementariedad
o incremento de la diversidad filogenÕtica para una regiµn determinada se puede calcular
aplicando las expresiones 1 µ 2, dependiendo de la cantidad de
informaciµn. En definitiva, todo depende del valor de conservaciµn que estemos
priorizando; si es la ![]() -diversidad
la expresiµn (4), si es la diversidad filogenÕtica, las fµrmulas (1 µ
2).
-diversidad
la expresiµn (4), si es la diversidad filogenÕtica, las fµrmulas (1 µ
2).
3. Las causas de la Biodiversidad
¢Cµmo se ha podido producir la diversidad de seres vivos que hoy observamos?, es decir, ¢cuÃles son los factores generadores de diversidad a lo largo del proceso evolutivo?. Posiblemente ningºn factor por sÚ solo, es capaz de explicar las causas de diversidad en una regiµn, siendo difÚcil la agrupaciµn de estos factores en conjuntos razonablemente homogÕneos. Tal vez por esta razµn, no disponemos aºn de una teorÚa general sobre las causas de la diversidad biolµgica y no es fÃcil discernir cuÃl es la contribuciµn diferencial de cada una de ellas en cada momento y lugar.
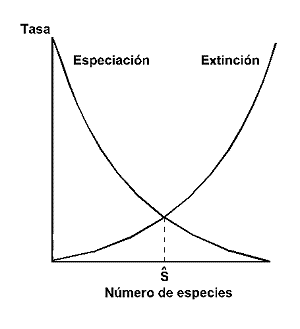
Una de las teorÚas ecolµgicas de mayor influencia en las ºltimas dÕcadas, el paradigma de MacArthur y Wilson (1963, 1967) sobre el equilibrio insular, postula que el nºmero de especies presentes en un lugar determinado, es el resultado de la tasa de generaciµn de especies (o de inmigraciµn en el caso de una isla) y la tasa de extinciµn (Figura 24).
Una primera divisiµn, un tanto acadÕmica, que permite una aproximaciµn bastante razonable consiste en considerar los factores intrÚnsecos, es decir, factores no generados por el medio y sÚ por las especies que integran la comunidad y los factores extrÚnsecos, es decir, aquellos que en ºltima instancia, dependen primordialmente del propio medio. Lobo & MartÚn Piera (1993) han desarrollo en extenso estos argumentos.
AdemÃs, la acumulaciµn incesante de datos de diversos organismos que colonizan las distintas Ãreas del planeta, va permitiendo establecer algunos patrones de variaciµn de la diversidad. Veamos los mÃs importantes.
3.1. Factores intrÚnsecos
El principio de Exclusiµn Competitiva establece que es imposible la supervivencia de dos especies en el mismo tiempo y lugar, si Õstas comparten idÕnticos recursos y variables ambientales. Ello viene a significar que no existen dos especies idÕnticas en sus adaptaciones, como consecuencia de la variabilidad inmensa de las condiciones particulares que han promovido la diferenciaciµn evolutiva de cada una. Aunque esta concepciµn ha de enfrentarse a la multiplicidad de conceptos de especie y con el desconocimiento de la influencia de las diferencias entre individuos de una misma especie, supone un punto de partida lµgico para la comprensiµn ecolµgica de la diversidad.
Todos los estudios teµricos y experimentales sugieren que la competencia entre dos especies por el mismo recurso, da lugar a la extinciµn de una de ellas pero la coexistencia tambiÕn es posible si el ambiente es heterogÕneo, si existe dispersiµn, si las especies se encuentran agregadas espacialmente, si se considera el papel de los depredadores, o bien, si se produce un desplazamiento de caracteres en alguna dimensiµn del nicho.
Surge asÚ el concepto de Similitud Limitante: existe un lÚmite en el nºmero de especies que pueden coexistir. De acuerdo con este principio es posible que la diversidad aumente si: 1) existe una mayor gama de recursos; 2) las especies poseen una amplitud de nicho pequeþa, es decir, estÃn especializadas en una o varias dimensiones del nicho y 3) la situaciµn contraria, las especies tienen mayor solapamiento de nicho.
Por otro lado, los predadores pueden ser selectivos y atacar a la especie o especies competitivamente superiores, permitiendo la coexistencia. TambiÕn pueden ejercer este efecto, si depredan sobre las presas mÃs abundantes en cada momento. La predaciµn puede permitir un grado mayor de solapamiento de nicho entre especies competitivas, al disminuir las densidades de poblaciµn y, por tanto, aumentar la capacidad de carga del medio. No obstante, la predaciµn puede acabar con la presa y, consiguientemente, con el recurso del propio depredador.
La Sucesiµn es otro factor generador de diversidad. La sucesiµn es el proceso temporal por el cual se modifica la composiciµn y las relaciones de abundancia entre las especies de una comunidad. Es un proceso tÚpicamente descrito en comunidades vegetales. En este caso las especies pioneras, es decir, las primeras en aparecer, modifican y promueven una mayor heterogeneidad en el medio de modo que crean nuevos recursos para el asentamiento de otras especies, incrementando la diversidad. Durante el proceso de colonizaciµn de una isla o del claro de un bosque tras un incendio, las especies van sucediÕndose unas a otras a lo largo del tiempo. Cuanto mÃs tiempo ha transcurrido, mayor es el nºmero de especies. Se acostumbra a considerar que los ecosistemas mÃs maduros son mÃs complejos, mÃs diversos. Por ello un territorio con parcelas en diferentes momentos de sucesiµn, albergarà mÃs especies que una Ãrea homogÕnea. Es importante seþalar que la Sucesiµn es tambiÕn un proceso de sustituciµn, es decir, no aumenta necesariamente la riqueza por adiciµn de especies.
3.2. Factores extrÚnsecos
Cuando hay poco alimento disponible, sµlo las especies selectivas pueden sobrevivir. Por el contrario, cuando abundan los recursos, encontramos muchas especies exigentes que estÃn especializadas en el consumo de un tipo particular de alimento. De este modo, los ecosistemas que tienen mayor producciµn de recursos, deberÚan tener mayor cantidad de especies. Las evidencias, sin embargo, no son concluyentes. Los ecµlogos han constatado en numerosas ocasiones que los ambientes muy productivos, tienen pocas especies: las tierras, los lagos o los rÚos sometidos a una intensa fertilizaciµn, aumentan su producciµn, pero disminuyen su diversidad. No hay respuesta convincente todavÚa. En realidad, la relaciµn entre productividad y biodiversidad es confusa. La mayor diversidad parece encontrarse en un umbral medio de productividad (Lobo, 1993).
Como un factor ligado a la productividad, debe considerarse la longitud de las cadenas trµficas. Ya se ha comentado anteriormente que la diversidad de una comunidad puede estar determinada por fuerzas estructuradoras diferenciales como la competencia y predaciµn, segºn sea la longitud de la cadena trµfica y el lugar que ocupe esa comunidad en dicha cadena trµfica.
Una zona con muchos tipos diferentes de suelos y superficies, tiene mÃs especies de plantas y un lugar con muchos tipos de Ãrboles alberga mÃs especies de aves e insectos que un monocultivo. Cuantos mÃs hÃbitats distintos tenga un territorio, mÃs especies diferentes puede albergar. El paisaje tiene muchas parcelas desiguales (parches) y esa heterogeneidad o mosaicismo ambiental, aumenta la diversidad; es la que Whittaker (1970 y 1975) denominµ Ô-diversidad; ya hemos hablado de ello. Esta podrÚa ser la causa de que haya mÃs especies en un territorio grande que en uno pequeþo.
Asimismo, la actividad de cualquier ser vivo crea heterogeneidad. AsÚ, los agujeros de picos picapinos sirven para que aniden otros animales, las hormigas transportan semillas de un lugar a otro y los ciervos, al matar Ãrboles cuando frotan su cornamenta contra la corteza, producen huecos en el bosque (Lobo, 1993).
|
Figura 25. Variaciµn del nºmero de especies de termitas de acuerdo con la latitud al norte del Ecuador. Datos de Sutton & Collins (1991). Modificado de Samways (1994). |
Figura 26. Variaciµn de la riqueza de plantas en montaþas del desierto de Arizona (Santa Catalina). Datos de Whittaker & Niering (1975). Modificado de Myers & Giller, (1988). |
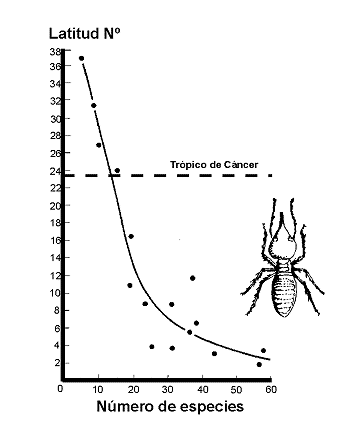
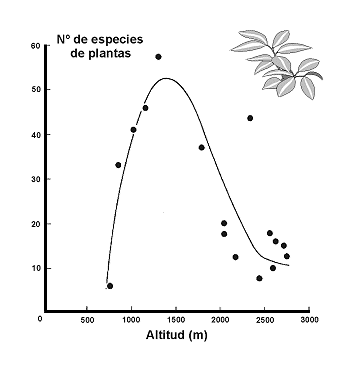
Una de las causas principales de que la diversidad sea menor en latitudes altas, es probablemente la inestabilidad climÃtica. En latitudes templadas la acusada estacionalidad promueve la diversidad. La cuestiµn, sin embargo, no està del todo clara. Los ambientes tropicales son mÃs estables y como es bien sabido alojan una gran diversidad (Figura 25). Lo que sucede es que ni todas las inestabilidades generan diversidad, ni los trµpicos son tan estables (Lobo, 1993).
La diversidad se modifica con muchos otros gradientes. Por ejemplo, varÚa con la altitud y con la profundidad. Las comunidades que viven en las zonas mÃs elevadas de las montaþas, estÃn muy aisladas y suelen habitar Ãreas muy pequeþas. Ello hace que la diversidad disminuya mÃs de lo normal con la altura. Todo parece indicar que el incremento de la diversidad se produce en rangos intermedios (Figura 26). Pero el patrµn, nuevamente no es generalizable (Figura 27).
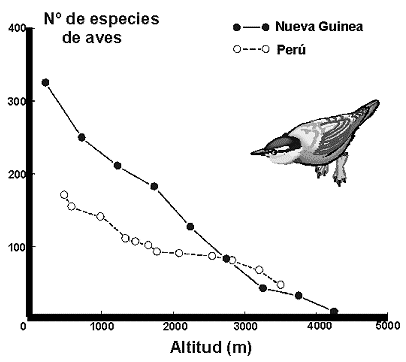
En mar abierto, la diversidad del plancton disminuye rÃpidamente con la profundidad. Se argumenta que la pÕrdida de luz es la responsable de este gradiente. Sin embargo, en los aþos sesenta se descubriµ que la diversidad del bentos es mÃxima a unos 2.000 m. de profundidad y despuÕs decae a medida que la luz y el alimento desaparecen (Figura 28); ¢cµmo puede ocurrir una cosa asÚ?. Tal vez ello sea debido al hecho de que estos ambientes han permanecido estables durante millones de aþos, permitiendo asÚ un incremento constante y paulatino de la Biodiversidad, como no ha sucedido en ningºn otro lugar del planeta.
En ocasiones la diversidad es producto de la acciµn combinada de varios de estos gradientes. AsÚ por ejemplo, la estacionalidad, la altitud y la orientaciµn, determinan conjuntamente la diversidad y la abundancia en las comunidades de escarabeidos coprµfagos.
En otras ocasiones la diversidad puede no estar relacionada con las caracterÚsticas actuales de un Ãrea, sino con las del pasado histµrico. Se piensa que cuanto mayor es la edad de una comunidad o un ecosistema, mayor es su diversidad (complejidad y estabilidad). Las especies que viven actualmente en el Norte de Europa, colonizaron este territorio tras la retirada de los ºltimos hielos hace 8.000 aþos. Durante el ºltimo medio millµn de aþos el clima, la fisiografÚa, la vegetaciµn y la fauna han experimentado cambios casi continuos a tasas muy superiores a las que prevalecieron durante los 200 millones de aþos precedentes.
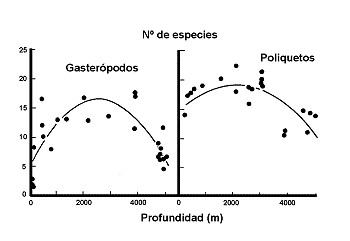
En ºltimo tÕrmino, los genes y los taxones representan los productos materiales de la evoluciµn, un proceso irreversible que ha tenido lugar a lo largo de 3.500 millones de aþos y que ha supuesto millones de eventos de diversificaciµn. Consecuentemente, los organismos representan diferentes combinaciones de sistemas genÕticos singulares y compartidos. Mientras que algunas partes del genoma tales como el RNA risobosomal son tan antiguas como la vida misma, a lo largo de la evoluciµn se han ido generando nuevos materiales genÕticos. Esta jerarquÚa histµrica de la vida, es la que los sistemÃticos intentan reflejar en las Clasificaciones, desde el Reino hasta las especies y las unidades infraespecÚficas.
Independientemente de que la jerarquÚa taxonµmica representa sµlo una fracciµn de la diversidad real (estimada entre 5-15 millones de especies; ver a continuaciµn Secciµn 4.1), la perspectiva histµrica de la diversidad es tambiÕn detectable a escala geogrÃfica, regional y local cuando analizamos las distribuciones geogrÃficas y ecolµgicas de genes, especies y taxones superiores. Ahora bien, asÚ como las genealogÚas son difÚciles de detectar debido a las homoplasias, los patrones geogrÃficos muy a menudo se complican por procesos estocÃsticos de dispersiones al azar o introducciones en hÃbitats no nativos. Sin embargo, podemos coincidir con LÕon Croizat en que la vida en la Tierra, muestra diferentes patrones que pueden ser descubiertos a travÕs de anÃlisis biogeogrÃficos. En suma, la diversidad biolµgica se caracteriza por el reemplazamiento espacial (‘turnover’) a todas las escalas geogrÃficas, por ello, la localizaciµn espacial de los componentes de la Biodiversidad, los seres vivos, es una tarea absolutamente crÚtica si pretendemos un diseþo efectivo de una red de conservaciµn in situ (Humphries, Williams & Vane-Wright, 1995).
Finalmente, no cabe duda que el hombre es un una especie que modifica rÃpidamente el medio, aumentado la inestabilidad de los ecosistemas y extinguiendo especies. Buena parte de los paisajes actuales de nuestro paÚs, no se entienden sin la intervenciµn humana. Por ejemplo, muchos pastizales ibÕricos, son el resultado de la actividad ganadera secular y por ello, una interrupciµn de la trashumancia podrÚa provocar el incremento de los arbustos y del fuego.
El lector interesado puede encontrar una excelente sÚntesis sobre la explicaciµn de la diversidad en Ricklefs (1995).
4. El Paradigma de la Biodiversidad: Los Insectos
4.1. El problema Taxonµmico
Ciertas medidas son consustanciales a nuestro conocimiento del Universo y sin ellas no se explica la imagen que nos hemos formado de Õl. ¢CuÃnto mide el diÃmetro de la Tierra?: 12.742 km.; ¢cuÃntas estrellas hay en la VÚa LÃctea?: aproximadamente 1011; ¢cuÃntos genes hay en un virus?: 10 en el fago X174; ¢cuÃl es la masa de un electrµn?: 9.1 x 10-28 gr; ¢cuÃntas especies de organismos habitan la Tierra?, no lo sabemos, ni siquiera el orden de magnitud (May, 1986, 1988, 1989, 1990 b, 1992; Hammond, 1995).
La cifra de Insectos se ha situado en torno a las 750.000 especies. Algunas estimaciones mÃs recientes elevan esta cifra a 1.112.000 (Samways, 1994) y, sin embargo, muchos entomµlogos hasta hace muy poco tiempo, habrÚan estimado que el nºmero de insectos no superarÚa el millµn y medio de especies. Williams (1964) doblµ esta estimaciµn a 3 millones de especies. Con estos datos Quintin Wheeler (1990) y el dibujante Francis Fawcett, han recreado un paisaje imaginario en el que cada grupo de organismos refleja las actuales estimas de diversidad (Figura 29). Sin embargo, Erwin (1982, 1983 a y b, 1988 y 1985) sugiriµ que en los bosques tropicales esta cifra se sitºa en un punto entre los 30 y los 50 millones de especies.
El tamaþo de los organismos representa el nºmero de especies en cada Taxon.
|
 |
Nºmero de especies descritas: |
|||
| TAXON | ESPECIES |
TAXON | ESPECIES |
| 1 Monera | 4,760 | 11 Mollusca | 50,000 |
| 2 Fungi | 46,983 | 12 Echinodermata | 6,100 |
| 3 Algae | 26,900 | 13 Insecta | 751,000 |
| 4 Plantas | 248,428 | 14 Artrµpodos no insectos | 123,161 |
| 5 Protozoos | 30,800 | ||
| 6 Porifera | 5,000 | 15 Pisces | 19,056 |
| 7 Coelenterata | 9,000 | 16 Amphibia | 4,184 |
| 8 Platyehelminthes | 12,200 | 17 Reptilia | 6,300 |
| 9 Nematoda | 12,000 | 19 Mammalia | 4,000 |
| Fuentes estadÚsticas: E. O. Wilson, Ed., 1988, Biodiversity, Nat. Acad. Sci. Press, Washington. | |||
| Figura 29. Paisaje imaginario creado por Wheeler (1980) y F. Fawcett, en el que el tamaþo de los organismos es proporcional a la diversidad taxonµmica de la Clase a que pertenece. Incluso con las estimaciones mÃs moderadas, el tamaþo proporcional de los insectos, representado en el grÃfico por un coleµptero, aºn deberÚa ser mayor. Modificado de Wheeler (1980). | |||
Esta estimaciµn causµ tal impacto que en los ºltimos cinco aþos ha sido uno de los temas mÃs recurrentes en la literatura entomolµgica. No faltan autores que consideran las estimaciones de Erwin ciertamente exageradas. AsÚ, Gaston (1991) argumenta que 10 millones de especies, es una estima mucho mÃs razonable de acuerdo con el conocimiento y experiencia de los diferentes especialistas que trabajan en todo el mundo. Erwin (1981) rebate los argumentos de Gaston, arguyendo que las estimas de los especialistas, no son un criterio cientÚfico sino un criterio de autoridad. La polÕmica aºn no ha terminado.
Es difÚcil sondear las implicaciones de estos nºmeros. Las cifras se disparan cuando las estimaciones se hacen en tÕrminos de individuos o biomasa (Wilson, 1991). AsÚ por ejemplo, tomando esta ºltima medida, se ha estimado que la biomasa de hormigas en los trµpicos, empequeþece la biomasa total de mamÚferos.
En todo caso, basÃndonos solamente en las especies descritas hasta la fecha, dos de cada cinco organismos vivos son un insecto. Ahora bien, si el nºmero de especies estimadas alcanza solamente los 10 millones de especies, es muy probable que esta proporciµn se incremente a 4 de cada 5 pero, probablemente, menos del 10 por ciento de los insectos han sido descritos formalmente con un nombre cientÚfico (Samways, 1993). Esto implica que cuando hablamos de extinciµn de especies y pÕrdida de Biodiversidad, estamos refiriÕndonos casi por completo a unos pocos megataxones: Insectos y otros invertebrados tales como, êcaros, AnÕlidos y Hongos, a pesar de su escasa presencia en las denominadas Listas Rojas (vÕase mÃs adelante Secciµn 4.3).
Es difÚcil documentar con precisiµn la extinciµn cuando muchas especies, particularmente de insectos, aºn no han sido descritas. Por otro lado, las tasas de extinciµn varÚan entre Ãreas geogrÃficas y tampoco son idÕnticas en todos los organismos (May et al., 1995). Las actuales estimaciones arrojan cifras verdaderamente alarmantes: Entre el 10 y el 25% de los seres vivos, podrÚan extinguirse en los prµximos 25-30 aþos (Raven, 1988); ¢a quÕ velocidad?. Con una estimaciµn tan moderada como 5 millones de seres vivos confinados en las Ãreas tropicales, Wilson (1988) popularizµ la cifra de 17,500 especies como la tasa anual de extinciµn en los bosques tropicales.
Hace ya una dÕcada la FAO estimµ que la destrucciµn anual del bosque lluvioso tropical es de 70,000 km2 anuales; un Ãrea casi equivalente a toda AndalucÚa, es decir, 40.5 Ha./3 minutos. Estimaciones mÃs recientes sitºan la tasa de deforestaciµn en el rango de 0.8-2% anual (Reid, 1992; Groom & Schumaker, 1993). Aplicando la relaciµn especies-Ãrea a la misma estimaciµn de 5 millones de especies, May et al. (1995) obtienen una extinciµn anual de 10,000-25,000 especies, o sea, 24-72 especies/dÚa, 1-3 especies/hora.
¢QuÕ significado tienen estas cifras en tÕrminos evolutivos?. Si no somos capaces de invertir la tendencia actual, la tasa actual de extinciµn podrÚa ser 400 veces superior a la del pasado geolµgico reciente. Los paleontµlogos estiman que la tasa promedio de extinciµn de especies es del 9%/millµn aþos, es decir, 0.000009 spp/aþo. Si aceptamos que la Biosfera reuniera una modestÚsima cifra de 2,000,000 especies, la tasa natural de extinciµn equivale a 0,18 spp/aþo (2.000.000 x 0.000009%); en cinco aþos: 0.18 x 5 aþos= 0,9 spp /5 aþos (. 1~5 aþos). Asumiendo como indican algunos paleontµlogos, que la estimaciµn es 10 veces superior (Raup, 1988), tendremos 10 spp/5 aþos, o bien 2 spp/aþo. Ahora bien, si el catÃlogo de la Biosfera ascendiera a 50,000,000 especies, como sugiriµ Erwin (1982, 1983 a y b 1985), el factor de proporcionalidad respecto al catÃlogo conocido es de 25, por tanto el factor de extinciµn natural deberÚa ser igualmente de 25. En consecuencia la tasa natural de extinciµn anual deberÚa ser 25 x 2 = 50 spp/aþo. Como la estima de extinciµn en valor absoluto es de 17,500-20,000 spp/aþo, resulta que la tasa real de extinciµn es 400 veces superior (50 x 400 = 20,000). El lector puede entretenerse en hacer el mismo ejercicio a partir de la cifra mÃs moderada de 10,000,000 de especies en toda la Biosfera. EncontrarÃ, en este caso, que el incremento de la tasa de destrucciµn (si sigue siendo cierto que desaparecen 20,000 spp/aþo) es 2,000 veces superior a lo que predicen los paleontµlogos; cifra que se sitºa en el rango de 1,000-10,000 estimado por Wilson (1988).
AsÚ las cosas nos enfrentamos a una paradoja y un dilema. La paradoja es que a medida que vamos incrementando nuestra capacidad de trabajo (descripciones taxonµmicas asistidas por ordenador, gestiµn informÃtica de enormes Bases de Datos faunÚsticos, esclarecimiento de patrones filogenÕticos y biogeogrÃficos, anÃlisis de procesos ecolµgicos, etc.), las especies estÃn desapareciendo incluso antes de que podamos llegar a conocerlas. Es fÃcil imaginar el dilema: ¢quÕ hacer?, ¢cuÃn rÃpido?. IntentarÕ contestar a esta cuestiµn como entomµlogo profesional valorando el trabajo realizado, la tarea pendiente y los recursos humanos disponibles.
Estos dilemas estÃn exacerbados por un complejo entramado de factores sociales, polÚticos y econµmicos alguno de los cuales, como la deforestaciµn, los acabo de mencionar. Otros en cambio, entran en el terreno de lo que solemos denominar polÚticas cientÚficas o sociologÚa de la ciencia, e incluso, en el juego de poder de los ‘lobbies’ cientÚficos; e.g.: Genoma Humano versus Inventario de la Biodiversidad.
|
Figura 30. Modelos de fluctuaciµn en poblaciones hipotÕticas. a: poblaciones fluctuantes prµximas a la mÃxima capacidad de carga del medio (K=20). Bajo este supuesto, las poblaciones estÃn afectadas por fluctuaciones ambientales dentro de un rango predominantemente favorable de condiciones microambientales. Este podrÚa ser el caso de una poblaciµn de insectos en un bosque tropical. b: La poblaciµn fluctºa en un amplio rango de densidades, entre K y L (lÚmite inferior), con extinciones locales ocasionales y reinvasiones. En este caso, la poblaciµn està afectada por cambios ambientales que originan incrementos y descensos demogrÃficos en la parte ‘intermedia’ del rango de condiciones microambientales. c: La poblaciµn fluctºa cerca del lÚmite demogrÃfico inferior (L=0) y la mayor parte del tiempo es ‘rara’, aunque està sujeta a explosiones ocasionales. En este caso, la poblaciµn existe a bajas densidades y està afectada por fluctuaciones que actºan sobre los individuos en el rango mÃs favorable del gradiente microambiental. Datos de Whittaker (1975). Modificado de Samways (1994). |
4.2. Las poblaciones de Insectos. La rareza de los Insectos
Las poblaciones de insectos difieren considerablemente en la constancia de sus niveles demogrÃficos y sus comunidades son muy dinÃmicas. Algunos son relativamente constantes de generaciµn en generaciµn, otros en cambio, varÚan frecuentemente y otros, en fin, se mantienen a niveles constantemente bajos y solo ocasionalmente son abundantes (Figura 30). La supervivencia de las poblaciones de insectos implica por tanto factores espaciales y temporales. Algunos estudios autoecolµgicos sobre mariposas, han mostrado que incluso especies muy localizadas y fuertemente amenazadas experimentan grandes variaciones de poblaciµn (Figura 31). Incluso cuando la poblaciµn es dividida en subunidades demogrÃficas, se puede constatar que algunas de estas subunidades pueden llegar a extinguirse, reaparecer y volver a extinguirse. Todos estos estudios demuestran que no es posible un buen conocimiento de las poblaciones de insectos sin una amplia perspectiva temporal. Ciertos rasgos del medio pueden parecer triviales en un aþo de observaciµn, al aþo siguiente igualmente banales con densidades muy diferentes y al siguiente mostrarse decisivos.
Por otro lado, Gaston & Lawton (1990) han seþalado que en insectos lo mÃs frecuente es una estrecha relaciµn entre abundancia local y rango geogrÃfico. Es decir, una especie localmente abundante, generalmente tiene un amplio rango geogrÃfico. No faltan sin embargo ejemplos en que no es posible establecer relaciµn entre abundancia y rango geogrÃfico.
Dos preguntas surgen inmediatamente, si las poblaciones de insectos fluctºan en el espacio y en el tiempo tan ampliamente, ¢cµmo adscribirlas a una categorÚa de rareza demogrÃfica, geogrÃfica o de hÃbitat? y ¢quÕ extensiµn debe tener el Ãrea geogrÃfica para tener una cierta garantÚa de Õxito en la conservaciµn?. DifÚciles cuestiones que nos reafirman nuevamente en la idea de que la conservaciµn de los insectos es una ficciµn sin un conocimiento bien fundamentado en estudios de fuerte dimensiµn temporal y espacial. Pero este tipo de estudios bÃsicos suponen un esfuerzo de inversiµn en recursos humanos y materiales que los gestores de la polÚtica cientÚfica y del Medio Ambiente, no siempre o casi nunca, estÃn dispuestos a financiar.
La rareza no sµlo està relacionada con la abundancia y el rango geogrÃfico sino tambiÕn con la tolerancia al biµtopo, es decir con su rango de euritopia o estenotopia; ya lo hemos visto (Figura 21).
Para acabar de complicar aºn mÃs las cosas la rareza en insectos tiene otras manifestaciones, no sµlo la demogrÃfica, tambiÕn un fuerte polimorfismo cromosµmico, enzimÃtico, ontogenÕtico y sexual asÚ como una gran variedad de ciclos biolµgicos alternativos y numerosas manifestaciones de mimetismo batesiano y m■lleriano (Samways, 1993). Todas estas manifestaciones de variabilidad requieren una diversidad de estrategias de conservaciµn. AsÚ por ejemplo, los diferentes estilos de ciclo biolµgico en insectos, son una variable importante a considerar desde el punto de vista de su preservaciµn. No podemos olvidar que muy a menudo, los distintos estados ontogenÕticos (especialmente en insectos holometÃbolos), se comportan como individuos muy diferentes desde el punto de vista ecolµgico, fisiolµgico, etolµgico y demogrÃfico. Conservar una especie de insecto holometÃbolo puede implicar la preservaciµn de hÃbitats muy diferentes dado que el adulto y las fases larvarias pueden vivir en medios diferentes, alimentarse de recursos igualmente diferentes y pueden sufrir presiones competitivas distintas. Por ejemplo los Escarabeidos adultos del gÕnero Cetonia son florÚcolas y palinµfagos sobre FagÃceas, CistÃceas, Compuestas, GramÚneas, etc.; las larvas, en cambio, viven a expensas de materia orgÃnica en descomposiciµn. Los Odonatos adultos son voladores y activos depredadores en el medio aÕreo; las larvas, sin embargo, son acuÃticas. Los escarabeidos coprµfagos adultos son de vida libre, compiten por el recurso trµfico y por el espacio para la nidificaciµn y pueden sufrir la depredaciµn; las larvas son endogeas, viven confinadas en habitÃculos-despensas o bolas-nido que les garantizan el alimento, la depredaciµn es menor pero se conocen casos de cleptoparasitismo.
4.3. La Conservaciµn de Insectos: Dilemas, Ficciones y ¢Soluciones?
Llegados a este punto cabrÚa preguntarse ¢por quÕ hay que conservar Insectos?. Hay varias Ãreas de interrelaciµn Hombre-Insecto, pero la importancia de esta interrelaciµn no ha sido aºn completamente establecida.
Aunque ºnicamente 200 especies de insectos son realmente plagas serias, su impacto es enorme. En realidad podemos responsabilizarlas de la pÕrdida de, al menos, el 30% de todas las cosechas mundiales. Actualmente se estima que el nºmero de plagas potenciales ronda los 10.000 especies de insectos. Incluso asÚ, esto es sµlo el 1% de las especies descritas. No menos agresivos parecen desde el punto de vista mÕdico y veterinario como vectores de graves enfermedades, algunas incluso mortales: malaria, dengue, leishmaniosis, enfermedad de Chagas, enfermedad del sueþo y un largo etc. No hay muchos motivos para la condescendencia.
AdemÃs de la precauciµn con este minºsculo nºmero de plagas, hay, en general, una gran dificultad para percibir la importancia de los Insectos en los procesos y dinÃmica de los ecosistemas. Sin embargo, cualquier plan global de conservaciµn debe tener en cuenta las necesidades de supervivencia en tÕrminos de funcionamiento ecolµgico, del conjunto de la biota terrestre (incluida nuestra propia especie) y los Insectos, son el componente cuantitativamente mÃs importante de los ecosistemas terrestres, aunque la funcionalidad de la mayorÚa de las especies, nos es aºn completamente desconocida.
AsÚ pues, si el Hombre està daþando la estructura y funcionamiento de la diversidad en la Biosfera y los Insectos son el componente mayoritario de esta Biodiversidad, estamos daþando, primordialmente, la diversidad de Insectos, antes incluso de que lleguemos a conocer cuÃles son sus ‘servicios ecolµgicos’. Esto no es chovinismo entomolµgico.
Con la excepciµn de unos pocos productos tales como la miel y algunos tintes, los Insectos poco tienen que ofrecer. Su pequeþo tamaþo, su esqueleto quitinoso y la gran variabilidad de sus poblaciones, les hace poco atractivos. Son sus ‘servicios’ ecolµgicos, su papel en la mayor parte de los ecosistemas terrestres como predadores, parasitoides, recicladores de nutrientes y materia orgÃnica, polinizadores, etc. (Samways, 1993), la razµn por la que deben ser conservados. Los hÃbitats y paisajes en los que ellos viven deben ser tambiÕn preservados y, en todo caso, utilizados de manera sustentable y duradera. AsÚ pues, los Insectos deben ser conservados per se, no por su valor econµmico que aºn està por establecer (Samways, 1993 y 1994).
Tendemos a considerar que la conservaciµn de especies se relaciona prima facies con las especies raras. Ya hemos visto las dificultades para aplicar este calificativo a los Insectos. Cambian de categorÚa incluso mÃs rÃpidamente que las plantas, debido a su variabilidad poblacional, particularmente en respuesta a los cambios del medio. Los Insectos ademÃs migran en respuesta a las modificaciones del paisaje, o en respuesta a cualquier modificaciµn biµtica (patµgenos) o abiµtica (clima). Las categorÚas de Rabinowtiz et al. (1986) son un marco conceptual muy ºtil en la conservaciµn de los Insectos. Sin embargo, la naturaleza dinÃmica de sus poblaciones, a corto plazo bajo la influencia de los paisajes antropizados y a largo plazo bajo la influencia de cambios geolµgicos o climÃticos, convierte a menudo en un artificio vacÚo de contenido biolµgico, la adscripciµn de las especies de insectos a las categorÚas de rareza.
Todo esto sin tener en cuenta el componente sociopolÚtico y hasta psicolµgico de la rareza. Una especie puede ser considerada rara en un paÚs y es completamente comºn en otro.
El conocimiento de la distribuciµn geogrÃfica de los insectos es esencial para tomar decisiones sobre su conservaciµn y preservaciµn. La escala espacial tiene un importante impacto en la percepciµn de la rareza, amenaza y estabilidad de las poblaciones. Los mapas de distribuciµn generalizada como los que estamos acostumbrados a ver en las guÚas de vertebrados, son de valor muy limitado en insectos con su pequeþo tamaþo y restricciones de hÃbitat y biµtopo. Muchos de estos mapas son mÃs potenciales que reales y, en el caso de los Insectos, a menudo reflejan la distribuciµn de los entomµlogos y no la del insecto en particular. En tÕrminos de urgencia el cartografiado de especies, paisajes, ecosistemas y tipos de biµtopos (Clark & Samways, 1992) y la predicciµn de las Ãreas potenciales de distribuciµn (Bartlein et al., 1986; Buckland & Elston, 1993; Scott et al., 1993; Huntley et al., 1995) mediante modelos de regresiµn (presencia/ausencia de especies en parcelas de territorio y las variables ambientales) (Smith, 1994) y/o la utilizaciµn de Sistemas de Informaciµn GeogrÃfica (SIG) (Jones et al., 1997; Skov & Borchsenius, 1997), son algunos de los objetivos mÃs inmediatos y realistas. Los SIG son una herramienta extremadamente valiosa para llevar a cabo esta tarea, eso sÚ, siempre y cuando seamos capaces de completar nuestras bases de datos biolµgicos. En nuestro paÚs, sin embargo, aºn no se dispone de un banco nacional de datos zoolµgicos, aunque muchos investigadores poseemos nuestras pequeþas bases de datos de trabajo.
Con todos estos datos en la cabeza, ¢quÕ significado biolµgico tienen los Libros Rojos y las Listas Rojas de Insectos?; muy poco. A pesar de su buena voluntad, frecuentemente, tales libros y listas son arbitrarias, subjetivas y estÃn fuertemente sesgadas desde un punto de vista antropocÕntrico hacia ciertos grupos ‘mÃs bonitos’, ‘mÃs grandes’ o ‘mÃs comºnmente conocidos’ tales como las mariposas o las libÕlulas.
La idea de que los Insectos merecen ser conservados no es nueva, pero es una idea que el hombre encuentra difÚcil aceptar. Estamos condicionados a creer que los Insectos deberÚan ser arrasados indiscriminadamente. Se interrelacionan muchas causas en esta percepciµn. Unas son de Úndole cientÚfica; ya las hemos comentado brevemente, otras son de Úndole psico-sociolµgica. El gran pºblico los repudia o mÃs a menudo los ignora, son molestos, daþinos o incluso vectores de enfermedades, a veces, mortales. En el fondo son nuestros mÃs directos competidores. Mantenemos una relaciµn esquizµtica con ellos que va desde su mitificaciµn como criaturas sagradas (vÕase MartÚn Piera o GrustÃn, en este mismo volumen) y la seducciµn por la belleza de sus formas y colorido, hasta el mÃs mortal de los odios: los insecticidas. Incluso desde un punto de vista estrictamente conservacionista, el tamaþo de los insectos es un elemento antropocÕntrico que sesga los criterios de conservaciµn a favor de otros organismos y tambiÕn dentro del propio mundo de los Insectos; los mÃs grandes y los mÃs bonitos. ¢A quiÕn se le ocurre conservar, por ejemplo, un pequeþo curculiµnido de 5 mm?, solamente a quien comprenda cuÃl es su ‘servicio’ ecolµgico. Lamentablemente, sabemos muy poco de la funcionalidad ecolµgica de la mayorÚa de los insectos; ya lo hemos comentado.
Desde que en 1835 Parnassius apollo fue declarada como especie protegida, unos pocos insectos han ido apareciendo tÚmidamente en la legislaciµn de algunos paÚses. CitarÕ sµlo algunas: En Inglaterra el ComitÕ para la Protecciµn de Insectos de la Real Sociedad Entomolµgica se remonta a 1925, en Francia la Oficina para la informaciµn eco-entomolµgica y en U.S.A. la Xerces Society fundada en 1971 (Samways, 1994). En los Convenios internacionales, sµlo algunos insectos aparecen recogidos en la Lista Roja de Animales amenazados de la IUCN (1990) y en los convenios de Berna (1971), Bonn (1979) y CITES (1973). Existen ademÃs algunas declaraciones de principios sobre el valor y Õtica de la conservaciµn de invertebrados, tales como la Carta de los Invertebrados Europeos adoptada por el Consejo de Ministros de la CEE en 1990 y en nuestro paÚs, el Manifiesto de Calpe sobre la gestiµn medioambiental y la conservaciµn de Artrµpodos (JimÕnez-Peydrµ & Marcos-GarcÚa, 1994; ver tambiÕn Galante, 1994 y Nieto-NafrÚa, 1994).
A pesar de este cºmulo de buenas intenciones ¢cuÃl es la realidad?. El Anexo IV de la Directiva Comunitaria 92/43 (1992), referido a las especies animales y vegetales que necesitan protecciµn en el Ãmbito de la CEE, recoge un total de 47 especies de Artrµpodos: 8 Coleµpteros, 23 Lepidµpteros, 12 Odonatos, 1 MÃntido, 2 Ortµpteros y un ArÃcnido (ver tambiÕn Rosas et al., 1992). La misma Directiva recoge mÃs de 160 especies de vertebrados. Con estos datos resulta evidente que los criterios de conservaciµn son de carÃcter claramente antropocÕntrico. Si tenemos en cuenta que el nºmero de especies que colonizan solamente la PenÚnsula IbÕrica asciende a 38.000, no parece exagerado afirmar que, hoy por hoy, la conservaciµn de insectos es una autÕntica ficciµn. ¢Quiere esto decir que no es posible?, en modo alguno, sµlo quiero decir que, simplemente, no se lleva a cabo.
Todo lo dicho hasta aquÚ arroja serias dudas sobre voluntad de conservaciµn de los Insectos y tambiÕn y aºn mÃs importante, sobre la validez de la aplicaciµn de los actuales criterios conservacionistas a estos organismos. Tal vez, ambos factores (falta de voluntad y de criterio), se interrelacionan mutuamente y son recÚprocamente sinÕrgicos. Es cierto que hace falta voluntad pero no es menos cierto y es aºn mÃs urgente, adoptar nuevos criterios que tomen en cuenta los peculiares desarrollos y biologÚa de los Insectos, sus grandes fluctuaciones demogrÃficas, sus cortos tiempos de generaciµn, sus adaptaciones a diferentes biµtopos. Todo sugiere que los Insectos se ajustan a dinÃmicas metapoblaciones con diferentes tasas de colonizaciµn entre subpoblaciones; distribuciones desiguales y fluctuaciones temporales tambiÕn desiguales, entre las subpoblaciones que colonizan diferentes biµtopos.
Teniendo en mente el gran impedimento taxonµmico y siendo el tiempo tan corto, hay una autÕntica necesidad de identificar Ãreas de mÃxima diversidad de insectos (‘Hot Spots’) y centros de endemicidad y/o rareza. Estas Ãreas crÚticas, son la ‘casa’ de especies taxonµmica y/o ecolµgicamente importantes y, tal vez, los centros de diversificaciµn actual y futura de las especies de insectos. Por estas razones Samways (1994) ha sugerido que hasta que no dispongamos de un Inventario razonablemente completo de estos organismos, deberÚamos utilizar el tÕrmino ‘comunidad amenazada’ en lugar de ‘especie amenazada’. Incluso en tÕrminos de organismos terrestres, Samways sugiere que aºn serÚa mÃs adecuado el tÕrmino de ‘paisaje amenazado’, incluyendo paisajes antropizados y mÃs o menos intactos.
4.4. Inventarios y Clasificaciones
Retomemos las dos preguntas iniciales: ¢quÕ hacer? ¢cuÃn rÃpido?. Cuatro frentes de ataque podrÚan aportar alguna soluciµn: i) acelerar el Inventario de la Diversidad OrgÃnica asÚ como nuestro conocimiento filogenÕtico; ii) estimar la diversidad a travÕs de grupos clave (indicadores) taxonµmicamente bien conocidos, en los diferentes ambientes, paisajes y ecosistemas, aplicando criterios ecolµgicos y sistemÃticos (Secciµn 2.1 y 2.2), a fin de establecer los puntos crÚticos o Ãreas de mÃxima diversidad; iii) investigar los procesos ecolµgicos relativos a la funcionalidad de los Insectos en los ecosistemas terrestres (Secciµn 2.2.) y iv) a tenor de nuestro desconocimiento general de la diversidad orgÃnica, su magnitud y sus causas, conservar y preservar mÃs de lo estimado, incluso a ciegas. Del segundo, tercer y cuarto puntos ya he hablado, me referirÕ ahora al primero.
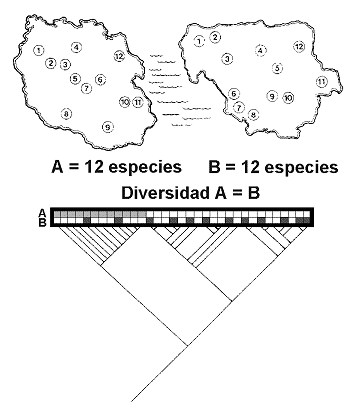
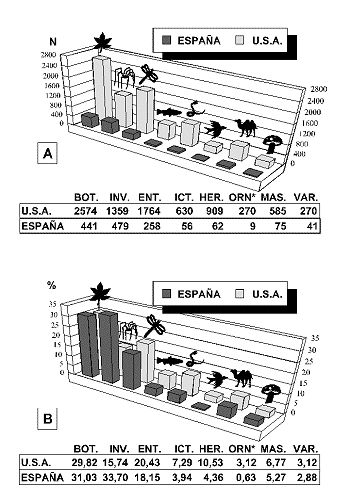
¢QuÕ es un Inventario de especies?. AquÚ surgen las primeras discrepancias no tanto sobre el quÕ es sino cµmo realizarlo; con quÕ criterios. En su mÚnima expresiµn, Janzen (1993) define un Inventario como una‘... base de datos taxonµmicamente limpia que indica, al menos, un punto geogrÃfico donde se encuentra una especie...’. A esta visiµn un poco simplista, Wheeler (1995) opone una concepciµn que separa lo que es el inventario propiamente dicho de la localizaciµn geogrÃfica. Wheeler define el Inventario del mundo orgÃnico como un Inventario Taxonµmico, esto es ‘... la exploraciµn, anÃlisis filogenÕtico y clasificaciµn predictiva de todas las especies de un taxon en todo el mundo...’ (Figura 32). AsÚ pues, una visiµn eminentemente pragmÃtica (Janzen) frente a una concepciµn rigurosamente cientÚfica (Wheeler); ambas sustentadas en el trabajo taxonµmico.
Llegados a este punto cabe preguntarse cuÃl es el estatus de la actual sistemÃtica de Insectos. Existe la opiniµn generalizada de que la SistemÃtica biolµgica es una actividad en franco declive. Veamos un par de ejemplos extremos: U.S.A y nuestro propio paÚs.
La figura 33 muestra las diferencias en valor absoluto y relativo entre ambas comunidades de sistemÃticos. Aunque las diferencias en valor absoluto son abismales, la tendencia de la distribuciµn por especialidades es, en tÕrminos porcentuales, comparable. En Estados Unidos los entomµlogos son cuantitativamente el segundo colectivo, tras los botÃnicos, en tanto que en nuestro paÚs, ocupamos la tercera posiciµn tras los invertebratµlogos. Estas cifras son mÃs fÃcilmente valorables con relaciµn al nºmero de organismos del que se ocupa cada colectivo de taxµnomos. Wheeler (1990) ha calculado la ratio taxµnomos/especies con los datos de la comunidad taxonµmica de EE.UU. y la diversidad mundial actualmente conocida. Obtiene unas cifras que para el caso de los entomµlogos sitºan esa ratio en 1:432. Con la estimaciµn de 10 millones esa ratio deberÚa ascender a r=1:5,669 y con las estimas de Terry Erwin (1982, 1983 a y b y 1985) alcanzarÚa r=1:28,345. Como tarea no està nada mal.
|
Figura 34. Ratio Taxµnomos/Organismos en la comunidad de sistemÃticos espaþoles. Bo.: BotÃnicos. In.: Invertebratµlogos. En.: Entomµlogos. Ic.: Ictiµlogos. He.: Herpetµlogos (anfibios y reptiles). Or.: Ornitµlogos. Ma.: Maztozoµlogos. Varios: Restantes phyla. Fuentes: ‘DIRTAX’ (GarcÚa-Valdecasas et al., 1994), Blanco & GonzÃlez (1992), datos inÕditos del Proyecto Fauna IbÕrica (Ramos, 1995) y estimaciµn de organismos vegetales a partir del Proyecto Flora IbÕrica (Castroviejo, 1995). Las cifras de organismos estÃn expresadas en centenares (x 100). |
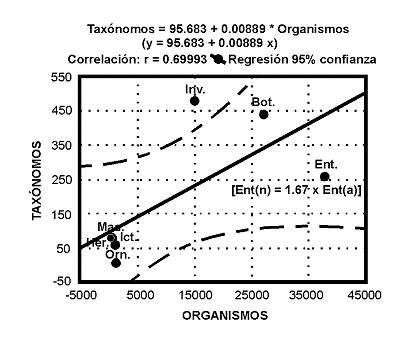 Figura 35. Regresiµn entre el nºmero de biosistemÃticos espaþoles, por especialidades, y magnitud taxonµmica de cada grupo en el Ãrea Úbero-balear. Bot.: BotÃnicos, Inv.: Invertebratµlogos. Ent.: Entomµlogos, Ent(n): Entomµlogos necesarios de acuerdo a la diversidad taxonµmica espaþola, Ent(a): Entomµlogos actualmente en activo. Ict.: Ictiµlogos. Her.: Herpetµlogos (anfibios y reptiles). Orn.: Ornitµlogos. Mas.: Maztozoµlogos. Fuentes: ‘DIRTAX’ (GarcÚa-Valdecasas et al., 1994), Blanco & GonzÃlez (1992), datos inÕditos del Proyecto Fauna IbÕrica (Ramos, 1995) y estimaciµn de organismos vegetales a partir del Proyecto Flora IbÕrica (Castroviejo, 1995). |
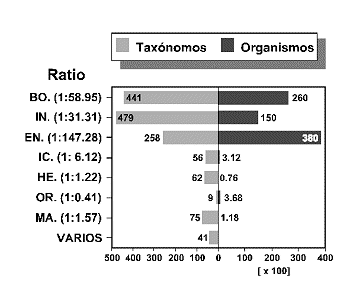
¢CuÃl es la situaciµn espaþola?. En nuestro paÚs con una estimaciµn de 38.000 spp. de insectos la ratio es 1:147.28. En relaciµn con las cifras de Wheeler la tarea se divide por 3 pero los taxµnomos se dividen por 7 (Figura 34). Es decir, el colectivo de entomµlogos que en nuestro paÚs se dedican a la taxonomÚa de entomofauna ibÕrica (la sÕptima parte de los que trabajan en USA), debe realizar un esfuerzo solamente tres veces inferior al ue deberÚan realizar todos los taxµnomos USA, dedicados a describir 750,000 especies de insectos. Como tarea tampoco està nada mal. En este sentido, resulta ilustrativo, la regresiµn entre ambos valores, taxµnomos y especies. La figura 35 indica claramente que en el momento actual, el colectivo de entomµlogos (258) resulta claramente insuficiente, estimÃndose que deberÚa incrementarse en un factor de 1.67 hasta alcanzar un nivel similar al de otras especialidades en nuestro paÚs, es decir, aproximadamente 430 especialistas.
Una de las propuestas mÃs audaces para finalizar en un plazo razonable de tiempo el inventario de la diversidad, fue la que lanzaron los comitÕs editoriales de las tres revistas sistemÃticas mÃs prestigiosas: ‘Cladistics’, ‘Systematic Botany’ y ‘Systematic Biology’. Esta iniciativa se conoce como ‘Systematic Agenda 2.000' (SA-200) (Blackmore & Cutter, 1996). Ahora bien, suponiendo que toda la Biota terrestre incluyera no mÃs de 5 millones de especies, al ritmo actual de descripciµn (£ 8,335 spp/aþo), tardarÚamos 6 siglos (600 aþos) en concluir el Inventario de la Diversidad OrgÃnica. Por consiguiente, la ºnica posibilidad de acabar dicho Inventario en 25 aþos, como proponen los promotores de la SA-2.000, serÚa multiplicar por 24 la tasa de descripciµn actual, pasando de las 8,335 ssp/aþo a las 200,000 spp/aþo (recordemos que el promedio anual de extinciµn de especies era de 167,000 spp/aþo). ¢Cµmo se hace esto? ¢multiplicando por 24 el nºmero de taxµnomos mundiales o exigiÕndoles 24 veces mÃs trabajo?. Yo no he encontrado nada en la interesante SA- 2.000 sobre esta cuestiµn. Antes al contrario, hoy sabemos que la tendencia general es mÃs bien la inversa, esto es, el abandono de la SistemÃtica biolµgica.
Si no parece posible o al menos es altamente improbable que seamos capaces de acelerar el Inventario de la Diversidad OrgÃnica, parece que la ºnica opciµn razonable es Conservar, es decir, mantener lo que tenemos con un uso ponderado o como ahora se dice, sustentable y en otros casos Preservar, o sea, mantener intactos algunos legados evolutivos singularmente valiosos (Samways, 1994).
1. Actualmente tenemos pocas estimaciones de la Biodiversidad y sabemos muy poco acerca de sus causas. Las fuerzas que los ecµlogos han propuesto, actºan a menudo en sentidos contrarios y es muy difÚcil emitir una opiniµn basada en hechos bien contrastados. Si supiÕramos cµmo se genera la Biodiversidad, podrÚamos establecer estrategias de protecciµn consecuentes, sustentadas en datos tangibles (MartÚn Piera & Lobo, 1992). Entre tanto, la actitud mÃs razonable es: i) ‘cubrirnos las espaldas’ protegiendo mÃs incluso de lo que nos parece necesario y, simultÃneamente, ii) promover la realizaciµn intensiva de estudios bÃsicos a medio y largo plazo. Entre Õstos, merecen especial atenciµn, los siguientes:
1.1. Descubrir, describir e inventariar la diversidad biolµgica con el contenido que Janzen (1993) y Wheeler (1995) dan a esta tarea y adquirir lo mÃs rÃpidamente posible el conocimiento filogenÕtico de las especies y grupos de especies, aunque sea un conocimiento in memorian, toda vez que la tasa de destrucciµn de hÃbitats y pÕrdida de especies, no tiene precedentes. Esta tarea, actualmente en franco declive en muchos paÚses, es especialmente acuciante en Insectos.
Es probable que la TaxonomÚa del futuro se nutrirà de una nueva generaciµn de tÕcnicos en identificaciµn (no necesariamente cientÚficos), expertos en anÃlisis de imagen, programaciµn y otras tÕcnicas informÃticas, que podrÃn escanear cientos de especies por dÚa e identificarlas mediante Sistemas Expertos (e.g.: ‘DELTA’ Dallwitz et al., 1993). Pero esto es sµlo tecnologÚa taxonµmica no investigaciµn taxonµmica. La TaxonomÚa como disciplina cientÚfica, habrà de seguir investigando la variabilidad inter e intrataxonµmica de los rasgos/atributos/caracteres a todos los niveles de la jerarquÚa taxonµmica y contrastando las hipµtesis evolutivas que en sÚ mismos establecen los propios taxones. No deberÚamos perder de vista esta distinciµn esencial entre Identificaciµn (TÕcnica) e Investigaciµn Taxonµmica (Ciencia).
1.2. Cartografiar los Puntos CrÚticos de la Biodiversidad, a escala global, con criterios sistemÃticos y ecolµgicos. Los primeros combinan la informaciµn filogenÕtica y biogeogrÃfica y, los segundos permiten establecer la distribuciµn potencial de los organismos analizando las variables ambientales que explican su distribuciµn geogrÃfica, o lo que es igual, el territorio que por sus caracterÚsticas ambientales o geogrÃficas debe contener la mayorÚa de las poblaciones de una especie. Para determinar el Ãrea potencial de distribuciµn de una especie se pueden utilizar modelos de regresiµn entre la presencia/ausencia de una especie en una parcela de territorio y los valores ambientales de dicha parcela, o bien, utilizar los denominados Sistemas de Informaciµn GeogrÃfica (SIG)
En el caso particular de los Insectos:
1.3. Investigar los factores (naturales o antrµpicos) que determinan las fluctuaciones de las poblaciones de insectos y el modo en que las comunidades responden a las perturbaciones de diversos agentes desestabilizadores.
2.1. El valor de los insectos como animales experimentales nos proporciona una magnÚfica ventana a la que asomarnos y comprender cµmo diferentes poblaciones, especies, ecosistemas y paisajes (antropizados o no), responden a cambios inevitables. Por otro lado, los insectos nos pueden enseþar mucho ya que configuran un mundo complejo que a lo largo de 350 millones de aþos de evoluciµn, ha ‘inventado’ casi todo. Existe una enorme diversidad de soluciones para resolver el mismo problema. Excepto el medio marino, han ocupado todos los rincones de la Tierra, adaptÃndose por ejemplo, a soportar temperaturas de 40 - 45¯C bajo cero y de 35 a 40 ¯C sobre cero. El abanico de posibilidades del estudio de los insectos es enorme: protecciµn contra la congelaciµn, envejecimiento, aprendizaje, estrategias reproductivas, etc. (BellÕs, 1995). No es casual el hecho de que el 32% de los artÚculos publicados en una de las mÃs prestigiosas revistas de EcologÚa (Journal of Animal Ecology), durante los ºltimos 61 aþos, hayan versado sobre insectos (Shorrocks, 1993).
3. Los Insectos como muchos otros microorganismos son un componente funcional vital de los ecosistemas casi completamente desconocido. A pesar de ello, siguen siendo objeto de escasa atenciµn en comparaciµn con otros organismos. Muy pocos insectos, incluso los incluidos en las Listas Rojas, estÃn individual y legalmente protegidos. A tenor del enorme impedimento taxonµmico en el estudio de los Insectos, se requiere una investigaciµn ingeniosa y aplicable. SerÚa muy importante que los entomµlogos hiciÕsemos especial hincapiÕ en el hecho de que la valoraciµn del potencial econµmico de las especies de insectos, està aºn por descubrir. Entre tanto, lo mÃs prudente es conservar y preservar tantos biµtopos, y paisajes como sea posible. Las Reservas y Parques Nacionales aunque no se designen especÚficamente para los Insectos, juegan un importante papel. Estas Reservas deberÚan ser tan grandes como fuera posible, a fin de alojar las ‘extravagancias’ de las poblaciones de insectos y evitar el peligro de extinciµn per se.
4. La variaciµn de las poblaciones de insectos es tan evidente en los trµpicos como en las Ãreas templadas. Pero la complejidad y variabilidad de tales poblaciones en el espacio y en el tiempo, dificulta extraordinariamente las predicciones en el balance demogrÃfico natalidad/mortalidad de los Insectos, incluso en las especies relativamente bien conocidas.
A pesar de ello, es preciso seguir en el tiempo la evoluciµn demogrÃfica de las poblaciones de insectos a lo largo de su rango geogrÃfico. Habida cuenta de que pequeþas poblaciones aisladas de insectos se extinguen con facilidad Ehrlich & Murphy (1987) han sugerido que serÚa deseable conservar un mÚnimo de amplitud metapoblacional.
Los individuos, las poblaciones, las especies y todas las interacciones, quedan protegidas de manera global con la conservaciµn del paisaje. A su vez, los Insectos pueden ser usados como un buen instrumento metodolµgico en la evaluaciµn de la conservaciµn de los paisajes (Samways, 1993). Los Insectos han demostrado ser buenos indicadores ambientales (Brown, 1991; Holloway & Stork, 1991; Pearson & Cassola, 1992; Halffter & Favila, 1993, Kremen et al., 1993; Favila & Halffter, 1997) aunque las evaluaciones con unas pocas especies proporcionan una ventana muy estrecha (Samways, 1994).
5. La naturaleza dinÃmica de las poblaciones de insectos, a corto plazo bajo la influencia de los paisajes antropizados y a largo plazo bajo la influencia de cambios geolµgicos o climÃticos, convierte a menudo en un artificio vacÚo de contenido biolµgico la adscripciµn de estos organismos a las categorÚas de rareza. No es de extraþar, por tanto, que los Libros Rojos y las Listas Rojas, tengan una fuerte carga subjetiva, arbitraria y hasta polÚtica, presentando a menudo un fuerte sesgo hacia ciertos grupos estÕticamente atractivos como las mariposas o las libÕlulas. Hoy por hoy, los Libros Rojos de insectos, son una simple guÚa, nada mÃs.
6. Es difÚcil imaginar un desarrollo social como el actual sin agredir al medio natural y de Õste, el elemento mÃs frÃgil, es la diversidad biolµgica. Sin embargo, si en la Õpoca postindustrial las sociedades humanas quieren ser dueþas de su propio destino, habrÃn de regular su actividad y crecimiento, aceptando que el techo de cualquier modelo de desarrollo, es el deterioro del legado mÃs importante de la evoluciµn biolµgica: La Biodiversidad (Halffter & Ezcurra, 1992). Tal vez hoy no lo acabamos de percibir con claridad, pero todo indica que en la conservaciµn y preservaciµn de la Biodiversidad, a escala global y su uso razonable (sustentable), asegura en ºltima instancia, nuestra propia supervivencia. Esta percepciµn global de la importancia de la Biodiversidad, trasciende uno de los conceptos mÃs tribales y autistas, pero tambiÕn mÃs querido, de nuestra sociedad: La SoberanÚa Nacional. Cada dÚa es mÃs evidente que los problemas medioambientales no pueden ser resueltos localmente. La diversidad biolµgica, no es una excepciµn. Los seres vivos y los ecosistemas en los que se estructuran, no entienden de reparto geopolÚtico. Por ello, es imprescindible derivar recursos econµmicos hacia los paÚses depositarios de las reservas de Biodiversidad.
7. Estas y otras muchas preocupaciones inspiraron la celebraciµn de la Cumbre de RÚo de Janeiro. Sin embargo, seamos realistas. Como ha seþalado di Castri (1993): ‘... La ºnica esperanza que surge del ‘EspÚritu de RÚo’ descansa casi exclusivamente en los ciudadanos, en la sociedad civil, en las fuerzas productivas y en las autÕnticas organizaciones no gubernamentales (muchas de las cuales no son sino la coartada de los gobiernos), quienes consolidan un frente de rechazo de lo inaceptable, rechazo a la creencia en documentos deliberadamente ambiguos, rechazo a legitimar lo anacrµnico y a las Instituciones estancadas (... algunas al borde de la ineptitud y obsolescencia...) ...’. MÃs adelante di Castri seþala que: ‘... hay demasiados mitos y palabras que han perdido su significado en los documentos de RÚo; es tal la ausencia de fundamentos cientÚficos, que el actual diÃlogo sµlo conduce al sÚndrome de la Torre de Babel, con nuevos gurºs, nuevos hechiceros y brujos. Los viejos demonios que compartimentan y sectorializan, nunca han estado tan presentes en las Instituciones (especialmente en los aspectos educativos y formativos) y entre los diferentes actores ...’. Di Castri, tampoco se olvida de la comunidad cientÚfica a la que acusa de ‘... entrar en el juego a fin de obtener financiaciµn para invertir en los mismos objetivos y las mismas intenciones que antes de RÚo, es decir, en proyectos que poco tienen que ver con los problemas reales ...’.
Personalmente creo que esta visiµn demoledora de di Castri, sobre los avances de la Cumbre de RÚo, se ajusta bastante bien a lo que es el dÚa a dÚa de la cuestiµn. No hay pues muchos motivos para el optimismo, para el idealismo; debemos esforzarnos en ser realistas. Sin embargo, justo es reconocer que el alegato de di Castri termina con una llamada al coraje y a la lucha contra este estado de cosas.
6. Una ºltima reflexiµn sobre los Insectos
La gente hallarà beneficio en la conservaciµn de los Insectos. No se puede vender bioÕtica a quien pasa hambre, pero se puede ilustrar el valor de los Insectos (Samways, 1994). Como indica este autor, la tarea de los entomµlogos es dar a conocer sus hallazgos cientÚficos a todos aquellos que hablan y pugnan por la preservaciµn de la Biodiversidad. Los hallazgos continuarÃn siendo de carÃcter taxonµmico descriptivo durante bastante tiempo, pero tambiÕn son hallazgos importantes el ºltimo enclave de un raro endemismo, el acusado perfil ecolµgico de una especie en el bosque lluvioso tropical, el desierto o un sistema de dunas, o la fragilidad de la fauna de una cueva singular µ, incluso, por quÕ no, algºn modelo sobre el impacto del calentamiento global. Toda esta informaciµn debe ser difundida a todos aquellos que quieran escuchar, polÚticos, gestores, ecologistas y pºblico en general y sobre todo, a las futuras generaciones.
Ackery, P. R. & Vane-Wright, R. Y., 1984. Milkweed Butterflies. British Museum (Natural History), London.
Alatalo, R. V., 1981. Problemas in the measurement of evennes in ecology. Oikos, 37: 199-204.
BaltanÃs, A., 1991. On the use of some methods for the estimation of species richness. Oikos, 65: 484-492.
Bartlein, P. J., Prentice, Y. C. & Webb, T., 1986. Climatic response surfaces from pollen data for some eastern North American taxa. Journal of Biogeography, 13: 35-37.
Baz, A., 1996. Ranking species and sites for butterfly conservation using presence-absence data in Central Spain. Nota lepidopterologica. Suppl., 2: 4-12.
Belles, X., 1995. Tendencias Actuales de la EntomologÚa IbÕrica. C. Albaladejo, X. BellÕs & F. MartÚn Piera. En: Avances en EntomologÚa IbÕrica, pp. 33-42. ComitÕ Editorial del VI Congreso IbÕrico de EntomologÚa (eds.). Museo Nacional de Ciencias Naturales (CSIC) y Universidad Autµnoma de Madrid. 502 pp.
Blackmore, S. & Cutler, D., (eds.) 1996. Systematic Agenda 2000. The challenge for Europe. Proceedings of a Workshop organised by the European Sciende Foundation Systematic Biology Network. Linnean Society Occasional Publication - N¤ 1. 105 pp.
Blanco, J. C. & GonzÃlez, J. L., 1992. Libro Rojo de los Vertebrados de Espaþa. Ministeriro de Agricultura, Pesca y Alimentaciµn. ICONA. Madrid. 714 pp.
Brooks, D. R. & McLennan, D. A., 1993. Phylogeny, Ecology , and Behaviour. A Research program in Comparative Biology. University of Chicago Press. Chicago. 434 pp.
Brooks, D. R. & McLennan, D. A., 1993. Historical ecology: Examining phylogenetic components of community evolution. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 267-280. University of Chicado Press. Chicago. 416 pp.
Brown, K., 1991. Conservation of Neotropical environments: Insects as indicators. En: Conservation of Insects and their Environments, pp. 349-404. N. M. Collins and J. A. Thomas (eds.). London, Academic Press.
Buckland, S. T. & D. A. Elston. 1993.- Empirical models for the spatial distribution of wildlife. Journal of Applied Ecology, 30: 478-495.
Caddle, J. E. & Greene, H. W., 1993. Phylogenetic patterns, Biogeography, and the Ecological structure of Neotropical Snake Assemblages. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 281-293. University of Chicado Press. Chicago. 416 pp.
Castroviejo, S., 1995. Flora IbÕrica. PolÚtica CientÚfica, C.I.C.Y.T., 44: 39-43.
Chapin III, F. S., Schulze, E.- D. & Mooney, H. A., 1992. Biodiversity and ecosystems processes. Trends in Ecology and Evolution, 7: 107-108.
Clark, T. E. & Samways, M. J., 1993. Ecological landscaping for conservation of macro-arthropod diversity in a southern hemisphere (Sourh African) urban botanic garden. En: Habitat Creation and Wildlife Conservation in Post-industrical and Urban Habitats. J. Rieley and S. Page eds. Packard. United Kingdom.
Cody, M. L., 1975. Towards a theory of continental species diversity bird distributions over Mediterranean habitat gradients. En: Ecology and Evolution of Communities M. L. Cody and J. M. Diamonds (eds.). Harward University Press, Cambridge, Massachussetts. pp. 214-257.
Colwell, R. & Coddington, J. A., 1994. Estimating terrestrial biodiversity through extrapolation. Philadelphia Transactions Royal Society. London, Ser. B., 345: 101-118.
Cornell, H. V. & Lawton, J. H., 1992. Species interactions, local and regional processes, and limits to the richness of ecological communities: A theoretical perspective. Journal of Animal Ecology, 61: 1-12.
Dallwitz, M. J., Paine, T. A., and Zurcher, E. J. (1993). `User’s Guide to the DELTA System: a General System for Processing Taxonomic Descriptions.’ 4th edition. (CSIRO Division of Entomology: Canberra.).
Darwin, Ch., 1859. On the Origin of Species by Natural Selection: or the Preservation of favoured races in the struggle for life. Ist Edition. London.
di Castri, F., 1993. One year after Rio: I’m guilty! BiologyInternational, 27: 1-2. Editorial.
Doube, B., 1990. A functional classification for analysis structure of dung beetle assemblages. Ecological Entomology, 20 (1): 90-97.
Ehrlich, P. R. & Murphy, D. D., 1987. Conservation lessons from long-term studies of checkersport butterflies. Conservation Biology, 1: 122-131.
Embley, T. M., Hirt, R. P. & Williams, D. M., 1996. Biodiversity at the molecular level: the domains, kingdoms and phyla of life. En: Biodiversity. Measurement and estimation. D. L. Hawksworth (ed.). pp. 21-33. Chapman & Hall. London. 140 pp.
Erwin, T. L., 1982. Tropical forests: their richness in Coleoptera and other arthropod species. Coleopterists Bulletin, 36: 74-75.
Erwin, T. L., 1983 a. Tropical forest canopies, the last biotic frontier. Bulletin of the Entomological Society of America, 29: 14-19.
Erwin, T. L., 1983 b. Beetles and other insects of tropical canopies at Manaus, Brazil, sampled by insecticidial fogging. Pages 59-75. En: Tropical rain forest: ecology and management S. L. Sutton, T. C. Whitmore, and A. C. Chadwick, editors. Blackwell Scientific Publications, Oxford U.K.
Erwin, T. L., 1985. Tambopata reserved zone, Madre de Dios, Peru: History and description of the reserve. Revista Peruana de Entomologia, 27: 1-8.
Erwin, T. L., 1988. The tropical forest canopy: the heart of biotic diversity. Pages 123-129. En: Biodiversity, E. O. Wilson, editor. National Academy Press, Washington, D. C.
Erwin, T. L., 1991. How many species are there?: Revisited. Conservation Biology, 5 (3): 330-333.
Faith, D. P., 1992 a. Conservation evaluation and phylogenetic diversity. Biological Conservation, 61: 1-10.
Faith, D. P., 1992 b. Systematics and conservation: on predicting the future diversity of subsets of taxa. Cladistics, 8: 361-373.
Faith, D. P., 1994. Phylogenetic diversity: a general framework for the prediction of feature diversity. Ed. P. L. Forey, C. J. Humphries & R. Y. Vane-Wright. pp.: 251-268. Systematics Association Special, vol. 50. Oxford University Press Inc., New York. 438 pp.
Faith, D. P., 1996. Phylogenetic pattern and the quantification of organismal biodiversity. En: Biodiversity. Measurement and estimation. D. L. Hawksworth (ed.), pp.: 45-58. Chpamand & Hall, 140 pp.
Farrell, B. D. & Mitter, Ch., 1993. Phylogenetic determinants of Insect/Plant community diversity. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 253-266. University of Chicado Press. Chicago. 416 pp.
Favila, M. E. & Halffter, G., 1997. The use of indicator groups for measuring biodiversity as related to community structure and function. Acta Zoologica Mexicana (n.s.), 72: 1-25.
Fisher, R. A., Corbet, A. S., & Williams, C. B., 1943. The relation between the number of species and the number of individuals in a random sample of animal population. Journal of Animal Ecology, 12: 42-58.
Galante, E., 1994. Los invertebrados, los grandes desconocidos en los programas de protecciµn medioambiental. En: Environmental Management and Artrhopod Conservation. pp. 75-88. Asociaciµn espaþola de EntomologÚa ed. 179 pp.
GarcÚa-Valdecasas, A., Bello, E. & Becerra, J. M., (eds.) 1994. DIRTAX: Directorio de Taxonµnomos Espaþoles. Graellsia Nºmero extraordinario. Museo Nacional de Ciencias Naturales. CSIC.. Madrid. 223 pp.
Gaston, K. J., 1991.- The magnitude of global insect species richness. Conservation Biology, 5 (3): 283-330.
Gaston, K. J. & Lawton, J. H., 1990. Effects of scale and habitat on the relationship between regional distribution and local abundance. Oikos, 58: 329-335.
Gauthier, J., Kluge, A. G. & Rowe, T., 1988. Amniote phylogeny and the importance of fossils. Cladistics, 4: 105-129.
Gibbons, D. W., Reid, J. B. & Chapman, R. A., 1993. The New Atlas of Breeding Birds in Britain and Ireland: 1988-91. Poyser, London.
Grombridge, B. (ed.) 1992. Global Biodiversity. Status of the Earth’s Living Resources. Chapman & Hall, London.
Groom, M. J. & Schumaker, N., 1993. Evaluating landscape change: patterns of worldwide deforestation and local fragmentation. En: Biotic interactions and global change. Kareiva, P. M., Kingsolver, J. G. & Huey, R. N., (eds.) pp. 24-44. Sinauer, Sunderland. Ma.
Halffter, G. & Ezcurra, E., 1991. ¢QuÕ es la Biodiversidad?. En: La Diversidad Biolµgica de IberoamÕrica, pp.3-24. Acta Zoologica Mexicana (n.s.). Volºmen especial de 1992. G. Halffter compilador. CYTED-D, Programa Iberoamericano de Ciencia y TecnologÚa para el Desarrollo. Mexico D. F. 389 pp.
Halffter, G. & Favila, M. E., 1993. The Scarabaeinae (Insecta: Coleoptera), an animal group for analyzing, inventoring and monitoring biodiversity in tropical rainforest and modified landscapes. Biology International, 27: 15-21.
Hammond, P., 1995. The Current Magnitud of Biodiversity En: Magnitud and Distribution of Biodiversity, Hawksworth, D. L. & Kalin-Arroyo, M. T. (Co-ordinat.). pp. 113-138. Chap. 3 of the Global Biodiversity Assessment. Heywood, V. H. & Watson, R. T. (eds.), United Nations Environment Program. Cambridge University Press. 1125.
Harper, J. L. & Hawksworth, D. L., 1996. Preface of Biodiversity. Measurement and Estimation. En: Biodiversity. Measurement and estimation. D. L. Hawksworth (ed.). pp. 5-12. Chapman & Hall. London. 140 pp.
Haydon, D., Radtkey, R. R. & Pianka, E. R., 1993. Experimental Biogeography: Interactions between Stochastic, Historical, and Ecological Processes in a model Archipielago. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 117-130. University of Chicado Press. Chicago. 416 pp.
Heltshe, J. F. & Forrester, N. E., 1983. Estimating species richness using the Jacnnife procedure. Biometrics, 39: 1-11.
Hill, M. O., 1973. Diversity and evennes: a unyfying notation and its consequences. Ecology, 54: 427-431.
Holloway, J. D. & Stork, N. E., 1991. The dimensions of biodiversity: The use of invertebrates as indicators of human impact. En: The Biodiversity of Microorganisms and Invertebrates: Its Role in _Suitable Agriculture, pp. 37-62. D. L. Hawksworth (ed.). CAB International, Wallingford, U.K.
Humphries, C. J., Williams, P. H. & Vane-Wright, R. I., 1995. Measuring Biodiversity value for conservation. Annual Review Ecology and Systematics, 26: 93-111.
Huntley, B. Berry, P. M., Cramer, W & McDonald, A. P., 1995. Modelling present and potential future ranges of some European higher plants using climate response surfaces. Journal of Biogeography, 22: 967-1001.
Hurlbert, S: H., 1978. The non-concept of species diversity: a critique and alternative parameters. Ecology, 59: 67-77.
IUCN, 1990. The 1990 IUCN Red List of Threatened Animals, IUCN Gland. Swuitherland, and Cambridge, United Kingdom.
Janzen D. H.,1993. What does tropical society want from the Taxonomists? En: Hymnoptera and Biodiversity. pp.: 295-307. J. Lasalle & Gauld, I. D., edit. C.A.B. International, 348 pp.
Jones, P. G., Sthephen. E., Joe Thome, B, Galwey, N. W., 1997.- The use of geographical information systems in biodiversity exploration and conservation. Biodiversity Letters, 6: 947-958.
JimÕnez-Peydrµ, R. Marcos-GarcÚa, M. A. (eds.) 1994. Environmental Management and Artrhopod Conservation. Asociaciµn espaþola de EntomologÚa ed. 179 pp.
Kikkawa, J. & Williams, E. E., 1971. Altitudinal distributio of land birds in New Guinea. Search, 2: 64-69.
Krebs, C. J., 1985. Ecology: The experimental analysis of Distribution and Abundance. Blackwell, Oxford.
Kremen, C., Colwell, R. K., Erwin, T. L., Murphy, D. D., Noss, R. F., & Saujayan, M. A., 1993. Terrestrial Arthropods assamblages; Their use in conservation planning. Conservation Biology, 7 (4): 796-808.
Lobo, J. M., 1993. La Base de la EcologÚa. Edit. Penthalon. Madrid. 155 pp.
Lobo, J. M., 1998. Influencias geogrÃficas, histµricas y filogenÕticas sobre la diversidad de las comunidades locales: una revisiµn y algunos ejemplos utilizando Scarabaeoidea coprµfagos (Coleoptera: Laparosticti). BoletÚn de la Asociaciµn espaþola de Entomologia, 21 (3-4): 15-31.
Lobo, J. M. & MartÚn Piera, F., 1993. Las causas de la Biodiversidad. Arbor, 145: 91-113.
Ludwig, J. A. & Reynolds, J. F., 1988. Statistical Ecology. A primer on methods and computing. John Wiley & Sons. New York. 337 pp.
MacArthur, R. H., 1957. On the relative abundance of bird species. Proceedings of the Natural Academy of Sciences, USA., 43: 293-295.
MacArthur, R. H., & Wilson, E. O., 1963. An equilibrium theory of insular zoogeography. Evolution, 17: 373-387.
MacArthur, R. H., & Wilson, E. O., 1967. The Theory of Island Biogeography. The Princeton Univeristy Press. Princeton.
McNaughton, S. J., 1995. Conservation goals and the configuration of biodiversity. En: Systematics and Conservation Evaluation. Ed. P. L. Forey, C. J. Humphries & R. Y. Vane-Wright. pp.: 41-62. Systematics Association Special, vol. 50. Oxford University Press Inc., New York. 438 pp.
McNeely, J. A., Miller, K. R., Reid, W. V., Mittermeier, R. A. & Werner, T. B., 1990. Conserving the world’s biological diveristy. IUCN, WRI, CI, WWF and World Bank, Washington, DC.
Magurran, A. E., 1988. Ecological Diversity and Its Measurement. Princeton University Press, New Jersey. 179 pp.
Margalef, R., 1958. Information theory in ecology. General Systematics, 3: 36-71.
MartÚn Piera, & FernÃndez Torres, A., 1996. Coleµpteros de la Sierra de Chiribiquete (CaquetÃ, Colombia). ELYTRON, 10: 23-50.
MartÚn Piera, F. & Lobo, J. M., 1992. La conservaciµn de la biodiversidad: ciencia y ficciµn. Eos, 68 (1): 91-92.
May, R., M., 1986. How many species are there?. Nature, 324: 514-515.
May, R., M., 1988. How many species are there on the earth?. Science, 241: 1441-1449.
May, R., M., 1989. How many species are there? En: The Fragile Environments. Friday, L. & Laskey, R., (eds.). Cambridge Univeristy Press, Cambridge, pp. 61-81.
May, R. M., 1990 a. Taxonomy as destiny. Nature, 347: 129-130.
May, R. M., 1990 b. How many species). Phylosophical transactions of the Royal Society, B330: 293-304.
May, R. M., 1992. Past efforts and future prospects towards understanding how many species there are. En: Biodiversity and Global Change. O. T. So.lbrig et al. (eds.) pp. 71-81. IUBS Monograph No. 8.
May, R. M., 1996. Conceptual aspects of the quantification of the extent of biological diversity. En: Biodiversity. Measurement and estimation. D. L. Hawksworth (ed.). pp. 13-20. Chapman & Hall. London. 140 pp.
May, R., M., Lawton, J. H. & Stork, N. E., 1995. Assesing extinction rates. En: Extinction Rates. Lawton, J. H. & May, R. M. (eds.) pp. 1-24. Oxford University Press. Oxford. 233 pp.
Mickevich, M. F. & Platnick, N. I., 1989. On the information content of classifications. Cladistics, 5: 33-47.
Myers, A. A. & Giller, P. S., (eds.) 1988. Analytical Biogeography. An integrated approach to the study of animal and plant distribution. Chapam & Hall. London. 578 pp.
Nieto-NafrÚa, J. M., 1994. Comentarios al ‘Manifiesto de Calpe’. En: Environmental Management and Artrhopod Conservation pp. 163-168. Asociaciµn espaþola de EntomologÚa ed. 179 pp.
Norse,E. A., Rosenbaum, K. L., Wilcove, D. S., Wilcox, B. A., Romme, W. H., Johnston, D. W. & Stout, M. L., 1986. Conserving biological diversity in our national forests. Washington, D. C.: The Wilderness Society.
Pearson, D. L., & Cassola, F., 1992. World-wide species richness patterns of tiger beetles (Coleoptera: Cicindelidae): indicator taxon for biodiversity and conservation studies. Conservation Biology, 6: 376-391.
Pearson, D. L. & Juliano, S. A., 1993. Evidence for the influence of Historical processes in Co-occurrence and diversity of tiger beetles. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 194-202. University of Chicado Press. Chicago. 416 pp.
Pielou, E. C., 1975. Ecological Diversity. John Wiley, New York.
Pielou, E. C., 1977. Mathematical Ecology. John Wiley, New York.
Prance, G. T., 1996. A comparison of the efficacy of higher taxa and species numbers in the assessment of biodiversity in the tropics. En: Biodiversity. Measurement and estimation. pp. 89-100. D. L. Hawksworth (ed.). Chapman & Hall, 140 pp.
Preston, F. W., 1948. The commonness, and rarity, of species. Ecology, 29: 254-283.
Rabinowitz, D., Cairns, S. & Dillon, Th., 1986. Seven forms of rarity and their frequency in the flora of British Isles. En: Conservation Biology. The Science of Scarcity and Diversity. Ed. Michael E. SoulÕ. pp. 182-204. Sinauers Associates, Inc. Massachusetts 584 pp.
Ramos, M. A., 1995. Fauna IbÕrica. PolÚtica CientÚfica, C.I.C.Y.T., 44: 35-38.
Raup, D. M., 1988. Diversity crises in the geological past. En: Biodiversity, E. O. Wilson editor, pp. 51-57. National Academy Press, Washington, D. C. 521 pp.
Raven, P. H., 1988. Our diminishing tropical forests. En: Biodiversity, E. O. Wilson editor, p. 119-122. National Academy Press, Washington, D. C. 521 pp.
Reid, W. V., 1992. How many species will there be?. En: Tropical deforestation and especies extinction. Whitmore, T. C. & Sayer, J. A., pp. 55-73. Chapman & Hall, London.
Ricklefs, R. E.., 1995. The distribution of biodiversity En: Magnitud and Distribution of Biodiversity, Hawksworth, D. L. & Kalin-Arroyo, M. T. (Co-ordinat.). pp. 139-173. Chap. 3 of the Global Biodiversity Assessment. Heywood, V. H. & Watson, R. T. (eds.), United Nations Environment Program. Cambridge University Press. 1125.
Ricklefs, R. E. & Schluter, D., 1993. Species Diversity: Regional and Historical influences. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 350-360. University of Chicado Press. Chicago. 416 pp.
Rosas, G., Ramos, M. A. & GarcÚa-Valdecasas, A., 1992.- Invertebrados espaþoles protegidos por Convenios Internacionales. ICONA. Ed. Madrid. 250 pp.
Rex, M. A., 1981. Community structure in the deep-sea benthos. Annual Review Ecology and Systematics, 12: 331-354.
Salt, G., 1979. A comment on the use of the term emergent properties. American Naturalist,113: 145-148.
Samways, M. J., 1990. Landforms and winter habitat refugia in the conservation of montane grassoppers in southern Africa. Conservation Biology, 4: 375-382.
Samways, M. J., 1983. Commnity structure of ants (Hymnoptera: Formicidae) in a series of habitats associated with citrus. Journal of Applied Ecology, 20: 833-847.
Samways, M. J., 1993. A spatial and process sub-regional framework for insect and biodiversity conservation research and management. En: Perspectives on Insect Conservation. Gaston, K. J., New, T. R. & Samways, M. J. (Eds.)., pp. 1-28. Intercept ltd., Andover, U.K., 250 pp.
Samways, M. J., 1994. Insect Conservation Biology. Chapman & Hall. London, 358 pp.
Scott, J. M., F. Davis, B. Csuti, R. Noss, B. Butterfield, C. Groves, H. Anderson, S. Caicco, F. D’Erchia, T. C. Edwards, J. Ulliman & R. G. Wright. 1993. Gap Analysis: A geographic approach to protection of biological diversity. Wildlife Monographs, 123: 1-41.
Shorrocks, B., 1993. Trends in Journal of Animal Ecology: 1932-1992. Journal of Animal Ecology, 62: 599-605.
Skov, F. & Borchsenius, F., 1997. Predicting plant species distributio patterns using simple climatic parameters: a case study of Ecuadorian palms. Ecography, 20: 347-355.
Smith, P. A., 1994. Autocorrelation in logistic regression modelling of species’ distributions. Global Ecology and Biogeography Letters, 4: 47-61.
Solbrig, O. T. (ed.), 1991 a. From genes to ecosystems: a research agenda for biodiversity. Cambridge, Massachusseetts,: International Union of Biological Sciences.
Solbrig, O. T., 1991 b. Biodiversity. A review of the scientific issues and a proposal for a collaborative program of research. MAB Digest 9, UNESCO.
Suttons, S. l. & Collins, N. M., 1991. Insect and tropical forest conservation. En: The Conservation of Insects and their habitats. Eds. N. M. Collins and J. A. Thomas. 15th Symposium of the Royal Entomological Society of London. AcademicPress, London. pp. 405-424.
Taylor, L. R., 1978. Bates, Williams, Hutchinson - a variety of diversities. En: Diversity of Insect Faunas: 9th Symposium of the Royal Entomological Society. Eds. L. A. Mound and N. Warloff, Blackwell, Osford, pp. 1-18.
Terborgh, J., 1977. Bird species diversity on Andean elevational gradient. Ecology, 58: 1007-1019.
Valkenbourgh, B. van & Janis, Ch. M., 1993. Historical diversity patterns in North American large herbivores and carnivores. En: Species Diversity in Ecological Communities. Historical and Geographical perspectives. Ricklefs, R. E. & Schluter, D. (eds.), pp. 230-340. University of Chicado Press. Chicago. 416 pp.
Vane-Wright, R. I., Humphries, C. J., Williams, P. H., 1991. What to protect?-Systematics and the Agony of a Choice. Biological Conservation, 55: 235-254.
Wheeler, Q. D., 1990. Insect diversity and cladistic constraints. Annals of the Entomological Society of America,83: 1031-1047.
Wheeler, Q. D., 1995. Systematic, the scientific basis for inventories of biodiversity. Biodiversity and Conservation, 4: 476-489.
Whittaker, R. H., 1960. Vegation of the Siskiyou Mountains, Oregon and California. Ecological Monographs, 30: 279-338.
Whittaker, R. H., 1970. Communities and Ecosystems. Macmillan, New York.
Whittaker, R. H., 1975. Communities and Ecosystems, 2ˆ ediciµn, Macmillan, New York.
Whittaker, R. H. & Niering, W. A., 1975. Vegetation of Santa Catalina Mountains, Arizona. V. Biomass, production and diversity along the elevational gradient. Ecology, 56: 771-790.
Williams, C. B., 1964. Patterns in the Balance of Nature and related problems in quantitative ecology. Academic Press London.
Williams, P. H., 1991. The bumble bees of the Kashmir Himalaya (Hymenoptera: Apidae, Bombini). Bulletin British Museum (Natural History), Entomology, 60: 1-204.
Williams, P. H., 1993. Measuring more of biodiversity for choosing conservation areas, using taxonomic relatedness. International Symposium on Biodiversity and Conservation (KEI) Korean Entomological Institute, Seoul, pp. 194-227 in Moon, T.-Y. (ed.)
Williams, P. H., 1995. Phylogenetic relationships among the bumble bees (Bombus Latr.): a re-appraisal of morphological evidence. Systematic Entomology, 19: 327-344.
Williams, P. H. & Gaston, K. J., 1994. Measuring more of biodiversity: can higher-taxon richness predict wholesale species richness? Biological Conservation, 67: 211-217.
Williams, P. H & Humphries, C. J., 1994. Biodiversity, taxonomic relatedness, and endemism in conservation. En: Systematics and Conservation Evaluation. Ed. P. L. Forey, C. J. Humphries & R. Y. Vane-Wright, pp. 269-287. The Systematics Association Special Vol. 50. Oxford Science Publications. 438 pp.
Williams, P. H. & Humphries, C. J., 1996. Comparing character diversity among biotas. En: Biodiversity. A Biology of Numbers and Difference, pp. 54-76. Ed. Kevin J. Gaston. Blackwell Science Ltd. Osford, 386 pp.
Williams, P. H., Vane-Wright, R. I. & Humphries, C. J., 1993. Measurign biodiversity for choosing convervation areas. En Hymenoptera and biodiversity. Ed. J. LaSalle & I. D. Gauld, pp. 309-328. CAB International, Wallingford, United Kingdom.
Williams, P. H., Humphries, C. J. & Gaston, K. J., 1994 a. Centres of seed-plants diversity: the family way. Proceedings of the Royal Entomological Society London. B 256: 67-70.
Williams, P. H., Gaston, K. J. & Humphries, C. J., 1994 b. Do conservationists and molecular biologists value differences between organisms in the same way?. Biodiversity Letters, 2: 67-78.
Williams, P. H., Gaston, K. J. & Humphries, C. J., 1997 a. Mapping biodiversity value worldwide: combining higher-taxon richness from different groups. Proceedings of the Royal Entomological Society, Biological Sciencies, 264: 141-148.
Williams, P. H., Vane-Wright, D., Humphries, C., Kitching, I., Lees, D., Araujo, M. & Gaston, K., 1997 b. Biodiversity. Measuring the variety of Nature & Selecting priority Areas for Conservation. Copyright Æ ‘The Natural History Museum Web Site’: http: //www.nhm.ac.uk/science/projects/worldmap
Williams, P. H., Gibbons, D., Margules, C., Rebelo, A., Humphries, C. J. & Pressey, R. A. A comparison of richness hotspots, rarity hotspots, and complementary areas for conserving diveristy using British birds. Conservation Biology; en prensa.
Williams, P. H, Prance, G. T., Humphries, C. J. & Edwards, K. S. Promise and problems in applying quantitative complementary areas for representing the diversity of some Neotropical plants (families Dichapetalaceae, Lecythidaceae, Caryocaraceae, Chrysoblanaceae and Proteaceae). Biological Journal of the Linnean Society; en prensa.
Wilson, E. O., 1988. The current state of biological diversity. En: Biodiversity, E. O. Wilson editor, pp. 3-18. National Academy Press, Washington, D. C. 521 pp.
Wilson, E. O., 1991. Ants. Wings, 16: 3-13.
1[ Teorema del LÚmite Central: Todas las variables estadÚsticas aditivas , tienen una distribuciµn Normal o Gausiana. En consecuencia, el logaritmo del nºmero de individuos tiene una variaciµn Normal o Gausiana a lo largo del tiempo.] [Volver]