ARACNET, 8 - Bol. S.E.A., nº 25 (1999) : 1-9.
Evolución de los Pachydeminae paleárticos (Coleoptera, Scarabaeoidea, Melolonthinae)
Doctoranda: Isabel SANMARTÍN BASTIDA
Director: Fermín MARTÍN PIERA
(1) Defendida en la Facultad de Ciencias Biológicas de la Universidad Complutense de Madrid, Departamento de Biología Animal (Entomología), el 18 de diciembre de 1998.
Dpto. Biodiversidad y B. Evolutiva (Entomología), Museo Nacional de Ciencias Naturales (C.S.I.C.), c/ José Gutiérrez Abascal, 2; 28006 - MADRID. e-mail: fermin@mncn.csic.es
ÍNDICE:
Antecedentes
Introducción
Objetivos
Material Estudiado
Metodología
Resumen de Resultados y Conclusiones
Literatura Citada
Antecedentes
En el tránsito al tercer milenio las sociedades industrializadas y en vías de desarrollo, han comenzado a percibir con claridad la necesidad de alcanzar un equilibrio entre las demandas de bienestar social y un crecimiento respetuoso con el medio natural, del cual dependen en última instancia, para su supervivencia individual y colectiva
En este contexto histórico, científico y socio-cultural, surge con fuerza un concepto unificador, tal vez un nuevo paradigma (Machado, 1998): La Biodiversidad. En los albores del siglo XXI, la conservación de la Biodiversidad en su sentido más holístico, dispara todas las alarmas de supervivencia de la sociedad civil y encauza todo el caudal científico de las tres disciplinas que a lo largo de los últimos ciento cincuenta años, se han consagrado al estudio de la Biosfera, sus elementos e interacciones, sus patrones de organización y procesos funcionales subyacentes en todos los niveles de organización, de las moléculas a los ecosistemas: La Biología Sistemática, la Ecología y la Biología Evolutiva. El nexo de unión de esta tríada natural (Edelstam et al., 1992) es la Biodiversidad, el gran diccionario de la Vida que se autoorganiza y enriquece mediante innumerables experimentos de evolución (Margalef, 1997).
La investigación desarrollada profundiza en uno de estos innumerables experimentos de evolución que contribuyeron a la diversificación de los coleópteros Melolóntidos, en un escenario histórico singular: el área mediterránea. Tras resolver numerosos problemas taxonómicos, se indagan los procesos históricos que pudieron dar origen a los actuales patrones filogenéticos y biogeográficos de los Pachydeminae paleárticos y, muy particularmente, los que colonizaron la cuenca mediterránea y la Península Ibérica. Se propone así, una teoría explicativa que da cuenta de la evolución espacial y temporal de estos coleópteros, en el marco de los actuales conocimientos paleogeográficos de la región paleártica occidental, constrastándola con las hipótesis biogeográfico-históricas que sugieren otros organismos.
Introducción
La diversidad entomológica de la Península Ibérica se estima en torno a las 38.000 especies (Martín Piera, 1997), lo que equivale a la quinta parte de la entomofauna mediterránea (Balleto & Casale, 1991). De éstas, aproximadamente 11.500 son coleópteros. Esta enorme diversidad apenas comienza a ser comprendida puesto que todavía un buen número de las familias de coleópteros íberobaleares, se encuentran en un nivel muy bajo de conocimiento taxonómico y faunístico.
La superfamilia Scarabaeoidea es una de las más diversas y mejor estudiadas dentro del orden Coleoptera. Su taxonomía y filogenia ha experimentado un gran avance en las últimas décadas gracias al empleo de nuevos caracteres de valor taxonómico y filogenético. Sin embargo, esto sólo es aplicable a los Scarabaeoidea Laparosticti, un vastísimo grupo de coleópteros descomponedores que explotan recursos efímeros tales como excrementos y carroña. Los Scarabaeoidea Pleurosticti, consumidores primarios, se encuentran todavía en un nivel elemental alfa-taxonómico de investigación. Salvo unas pocas especies responsables de plagas de cultivos, muy poco se sabe sobre la biología, dinámica poblacional, adaptaciones ecológicas y comportamiento de estos coleópteros.
|
|

La presente tesis aborda el estudio taxonómico, filogenético y biogeográfico-histórico de un grupo apenas conocido de Scarabaeoidea Pleurosticti: la subfamilia Pachydeminae (Melolonthidae) (fig. 1). Los Pachydeminae forman una parte muy importante de la fauna edáfica que coloniza el horizonte radicular: la Rizosfera. Sin embargo, en comparación con otros grupos de Pleurosticti como los Melolonthinae o los Rutelinae, la taxonomía de los Pachydeminae ha sido muy poco estudiada (Baraud, 1992).
Esta investigación se circunscribe al área paleártica, donde la subfamilia reúne 16 géneros (Baraud, 1992): Peritryssus Reitter, 1918; Hemictenius Reitter, 1897; Pachydema Castelnau, 1832; Tanyproctus Faldermann, 1835; Atanyproctus Petrovitz, 1955; Leptochristina Baraud & Branco, 1991; Elaphocera Gené, 1836; Ceramida Baraud, 1987; Pachydemocera Reitter, 1902; Brenskiella Berg, 1898; Europtron Marseul, 1867; Alaia Petrovitz, 1980; Tanyproctoides Petrovitz 1971; Pseudopachydema Balthasar, 1930; Otoclinius Brenske, 1896; Kryzhanovskia Nikolajev & Kabakov, 1977.
En la Península Ibérica, la subfamilia Pachydeminae está representada por dos géneros, Ceramida Baraud, 1987 y Elaphocera Gené, 1836, con un total de 60 especies descritas. La mayor parte presentan un acusado carácter endémico, probablemente debido a la condición braquíptero-áptera de las hembras, lo que ha determinado una escasa movilidad y rangos geográficos muy reducidos. El género Ceramida es prácticamente endémico de la Península, el 90% de sus especies son ibéricas, este porcentaje desciende al 50% en el género Elaphocera. Estos géneros se caracterizan por su gran dificultad taxonómica (Báguena, 1955; Branco, 1981; Baraud, 1992). Las especies son muy parecidas en su morfología por lo que es difícil encontrar rasgos diagnósticos de valor taxonómico. Asimismo, la escasez de material en las colecciones, dificulta la valoración de la variabilidad intraespecífica de tales rasgos. Esto ha desembocado en la frecuente descripción de nuevas especies, basadas en uno o muy pocos ejemplares (López-Colón, 1986; Blanco-Villero, 1988; Ruiz & Avila, 1995). En Ceramida, la falta de una taxonomía clara ha dificultado la identificación de las especies plaga y de su posible control biológico (Alvarado et al., 1996).
De acuerdo con las ideas precedentes, el interés de esta investigación radicaba en tres aspectos distintos:
a) Taxonómico: Los problemas taxonómicos señalados, han propiciado una taxonomía inestable y poco fiable. Se han descrito numerosos géneros monoespecíficos de posición taxonómica dudosa (Piatella & Sabatinelli, 1996) y varios de los géneros más grandes, se han definido por caracteres de valor taxonómico poco contrastado (Tanyproctus, Pachydema). Exceptuando la monografía de Jacques Baraud (1992), limitada a la región paleártica occidental, no existe una obra de conjunto del grupo sino tan sólo trabajos parciales en los que se revisa un género o un grupo de especies (Branco, 1981; Nikolaev, 1987; Baraud, 1991; Baraud & Branco, 1991, etc.). Además, la identificación taxonómica descansa casi exclusivamente en los caracteres de la morfología externa, que han demostrado ser insuficientes para la caracterización de los grupos.
b) Filogenético: No existe ningún estudio sobre las relaciones filogenéticas de la subfamilia Pachydeminae en este área. Una clasificación de la subfamilia basada en la inferencia filogenética, permitiría superar la inestabilidad taxonómica actual y comprobar el carácter monofilético de los géneros.
c) Biogeográfico: Los Pachydeminae reúnen tres condiciones que les convierten en una herramienta ideal para contrastar hipótesis biogeográficas de carácter histórico: gran diversidad taxonómica, alta endemicidad y una distribución geográfica heterogénea. Se reconocen 68 géneros en todo el mundo y su distribución disyunta sugiere un origen muy antiguo en el paleocontinente Gondwana. Se conocen Pachydeminae en el Sur de África, regiones de Australia y de las Penínsulas Malaya e Indostánica y en el continente americano: Perú, Argentina, Chile, sur de México y extremo oeste de los EE.UU. (California). En el área paleártica presentan una distribución anfimediterránea meridional: Magreb, Península Ibérica e islas del Mediterráneo occidental, en el oeste, y Balcanes, Anatolia y Asia Menor en el Mediterráneo oriental; no se conocen Pachydeminae en la península italiana y mediodía francés. Algunos géneros colonizan el Cáucaso y la llanura aralo-cáspica.
A diferencia de los Melolonthinae (Melolontha Fabricius, 1775, Polyphylla Harris, 1842, Anoxia Castelanu, 1832), los Pachydeminae carecían de interés económico (Balachowsky, 1962) pues hasta muy recientemente, no se conocía ninguna especie que representara un problema agronómico. Tal vez, esta es una de las causas por las que apenas se tienen datos sobre la biología de las especies y sólo se han descrito estados larvarios en dos géneros: Hemictenius y Tanyproctus (Medvedev, 1960; Nikolaev, 1987). Sin embargo, en 1993 se denunció la existencia de una nueva plaga de los cultivos de olivo y vid en el Sur de España, ocasionada por larvas del género Ceramida (Alvarado et al., 1995; Serrano et al., 1996). Esta plaga, producida por cambios en las técnicas de cultivo, demuestra la capacidad de las especies de Pachydeminae para convertirse en plagas potenciales.
Objetivos
El objetivo central de la Tesis es el desarrollo de una teoría explicativa sobre la evolución espacial y temporal de los Pachydeminae en el área paleártica. Para la consecución de este objetivo, el curso de la investigación siguió tres fases:
I. Taxonomía:
|
|
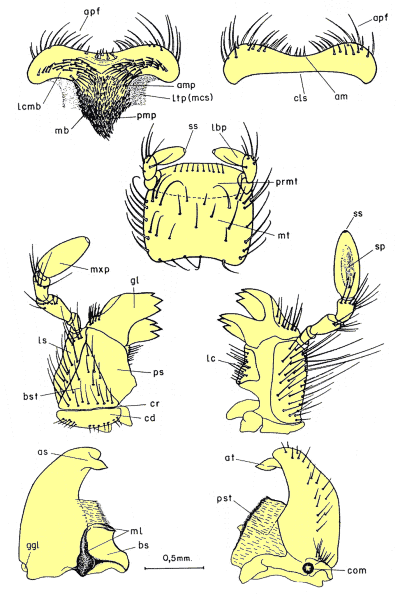
i) Búsqueda de nuevos caracteres de valor taxonómico que pudieran ser utilizados para aclarar la taxonomía del grupo y, posteriormente, como hipótesis de homología en la reconstrucción filogenética. Para ello, se revisó el valor taxonómico de los caracteres de morfología externa tradicionalmente utilizados en la taxonomía del grupo, comprobando su variabilidad intraespecífica y se exploraron tres nuevas homologías no estudiadas en Pachydeminae: el aparato bucal (fig. 2), la anatomía alar (fig. 3) y la genitalia de ambos sexos (figs. 4, 5 y 6).
ii) Aplicación de los nuevos caracteres de valor taxonómico para resolver la taxonomía específica de los dos géneros ibéricos de Pachydeminae: Elaphocera y Ceramida. En este último género, se estudió también la biología de las especies plaga de los cultivos y se describió la larva de tercera edad de Ceramida cobosi.
II. Filogenia:
i) Reconstrucción de las relaciones filogenéticas de los géneros paleárticos de Pachydeminae, con el fin de comprobar su carácter monofilético y establecer una nueva clasificación natural de la subfamilia.
ii) Análisis de las relaciones filogenéticas de las especies de los géneros ibéricos Ceramida y Elaphocera. Esto ha permitido establecer una nueva clasificación que resuelve la inestabilidad taxonómica actual.
III. Biogeografía Histórica:
Formulación de la primera hipótesis sobre la historia biogeográfica de los Pachydeminae paleárticos. De acuerdo con los datos paleogeográficos, se exploraron hipótesis alternativas de evolución espacial en diferentes escenarios biogeográficos históricos, capaces de explicar los actuales patrones de distribución geográfica de la subfamilia en el área paleártica. Posteriormente, se establecieron las vías de colonización de los géneros ibéricos de Pachydeminae, Ceramida y Elaphocera, en la península Ibérica y en el área mediterránea.
|
Figura 3. Estructura del ala en Pachydeminae: 1) Género Pachydemocera; 2) Género Europtron. AA (1+2, 3+4): Vena anal anterior (primera y segunda división). AP: Anal posterior; CuP: Cubital posterior |
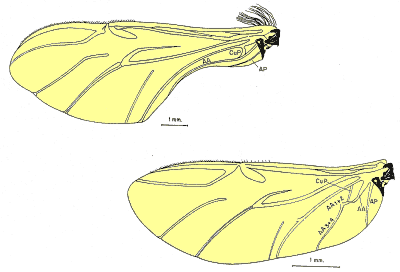
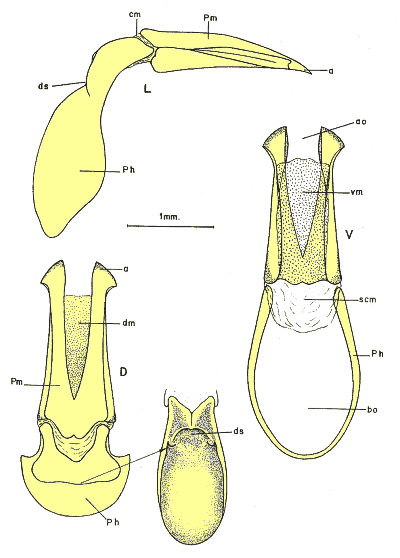 Figura 4. Tegmen en visión lateral (L), dorsal (D) y ventral (V) de Pachydema a: ápice de los parámeros; ao: ostium apical; bo: ostium basal; cm: membrana conectiva; dm: membrana dorsal de los parámeros; ds: estrangulación dorsal de la falobase; Ph: falobase; Pm: parámeros; scm: segunda membrana conectiva; membrana ventral de los parámeros. |
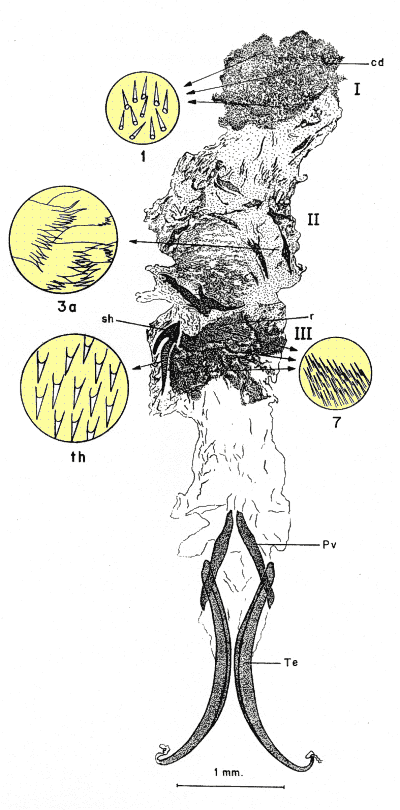
Figura 5. Endofalo de Pachydema hirticollis (Fabricius, 1787) mostrando la división en bandas o áreas de sedas de distinta morfología: I: área distal; II: área media; III: área basal. Detalles de las sedas, 1: sedas en arpón; 3a: sedas digitiformes y aplanadas, anchas; 7: ráspula de sedas múltiples. Otras estructuras, cd: divertículo caudal del endofalo; Pv: pieza en "V"; r: ráspula; sh: estructura esclerotizada del endofalo; Te: temonas; th: detalle de los dientes de la estructura esclerotizada.
|
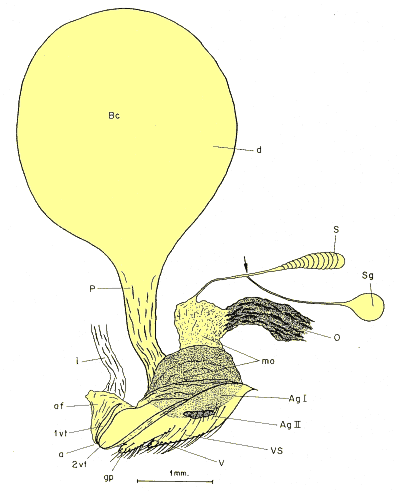 Figura 6. Genitalia femenina de Ceramida tangeriana (Kraatz, 1882). Vista lateral. a: ano; af: pliegue anal; AgI, AgII: glándulas accesorias; Bc: bolsa coplatriz; d: divertículo de la bolsa copulatriz; gp: palpo genital; i: intestino: recto; P: pedúnculo; S: espermateca o receptáculo seminal; Sg: glándula de la espermateca; v: vagina; 1vt, 2vt: terguitos vestigiales; vs: esternitos vestigiales. |
Material Estudiado
En la revisión de los Pachydeminae, se incluyeron los 16 géneros de que consta la subfamilia en el área paleártica. Solamente en el caso de los géneros monoespecíficos Pseudopahydema y Kryzhanovskya no se pudieron estudiar los tipos, único material conocido, por lo que los caracteres se tomaron de la descripción original.
Se estudió una muestra representativa de las especies teniendo en cuenta tres criterios: i) representación de la variabilidad geográfica del género, de forma que estuvieran representadas todas las áreas de distribución por, al menos, una especie; ii) representación de la variabilidad morfológica. Esto es especialmente importante en el caso de los géneros con gran número de especies tales como Elapocera, Ceramida, Tanyproctus, Pachydema y Hemictenius, donde la morfología externa es muy variable; iii) representación de la variabilidad taxonómica, incluyendo al menos, una especie de cada uno de los subgéneros o grupos de especies que otros autores han distinguido en géneros muy polimorfos tales como Pachydema y Tanyproctus (Baraud, 1985).
En el caso de los géneros ibéricos Elaphocera y Ceramida, se estudiaron todas las especies conocidas. Se revisaron además la casi totalidad de las sinonimias propuestas anteriormente, incluyendo las series típicas.
La Tabla 1 resume todo el material analizado. Se ha estudiado el 86% de las especies de Pachydeminae en el área paleártica, por lo que se considera que este trabajo representa adecuadamente su diversidad en dicha área. En total, se examinaron más de 2000 ejemplares.
Las colecciones institucionales fueron las siguientes: Museo de Ciencias Naturales de Madrid (MNCN); Museo Nacional de Historia Natural de Paris (MNHN); Museo de Historia Natural de Hungría (HNHM); Museo de Historia Natural de Ginebra (MHNG); Museo de Historia Natural de Viena (MHNV); Museo Nacional de Historia Natural de Leiden (NNM); Museo de la Universidad Humtboldt de Berlín (MUHB); Instituto de Entomología Alemán de Eberswalde (DEI); Smithsonian Institution (SI); Museo Americano de Historia Natural (AMNH); Universidad de Amsterdam (UvA); Senckemberg Instituto de Frankfurt (FSI); Museo de Zoología Comparada de Harvard (MCZ); Museo de Historia Natural de la Universidad de Coimbra (MHNCo).
También se estudiaron tres colecciones particulares: Colección Artur Serrano; Colección J. I. López-Colón, y Colección J. M. Ruiz.
Tabla 1:
Géneros paleárticos de Pachydeminae. Número de especies estudiadas dentro de cada género y porcentaje respecto al total de especies
|
Géneros |
Nº spp. estudiadas |
Nº spp. total por género y fuente |
% s/. Total |
|
Alaia Petrovitz, 1980 |
1 |
1 (Petrovitz, 1980) |
100 % |
|
Atanyproctus Petrovitz, 1955 |
4 |
6 (Petrovitz, 1980) |
67 % |
|
Brenskiella Berg, 1898 |
1 |
1 (Baraud, 1992) |
100 % |
|
Ceramida Baraud, 1987 |
18 |
18 (Baraud, 1992, y trabajos posteriores) |
100 % |
|
Elaphocera Gené, 1836 |
43 |
43 (Baraud, 1992 y trabajos posteriores) |
100 % |
|
Europtron Marseul, 1867 |
2 |
2 (Baraud, 1985) |
100 % |
|
Hemictenius Reitter, 1902 |
11 |
24 (Nikolaev, 1987) |
46 % |
|
Kryzhanovskia Nikolaev & Kabakov, 1977 |
1 |
1 (Nikolaev & Kabakov, 1977) |
100 % |
|
Leptochristina Branco & Baraud, 1991 |
4 |
4 (Branco & Baraud, 1991) |
100 % |
|
Otoclinius Brenske, 1896 |
4 |
4 (Baraud, 1991) |
100 % |
|
Pachydema Castelnau, 1832 |
23 |
77 (Baraud, 1985) |
30 % |
|
Pachydemocera Reitter, 1902 |
1 |
1 (Baraud, 1992) |
100 % |
|
Peritryssus Reitter, 1918 |
1 |
1 (Baraud, 1992) |
100 % |
|
Pseudopachydema Balthasar, 1930 |
1 |
1 (Baraud, 1992) |
100 % |
|
Tanyproctoides Petrovitz, 1971 |
1 |
1 (Petrovitz, 1971) |
100 % |
|
Tanyproctus Faldermann, 1835 |
18 |
50 (Reitter, 1902, Nikolaev, 1987, Zool. Record) |
36 % |
Listado de especies estudiadas por género
|
• Alaia Petrovitz, 1980 |
|
A. sexdentata Petrovitz, 1980 |
|
• Atanyproctus Petrovitz, 1955 |
|
A. miksici Petrovitz, 1965 |
|
A. simplicitarsis Petrovitz, 1955 |
|
A. ernae Petrovitz, 1980 |
|
A. afghanus Petrovitz, 1968 |
|
• Brenskiella Berg, 1898 |
|
B. flavomicans Berg, 1898 |
|
• Ceramida Baraud, 1987 |
|
C. abderramani (Escalera, 1923) |
|
C. baraudi (Branco, 1981) |
|
C. bedeaui (Erichson, 1840) |
|
C. brancoi (Baraud, 1975) |
|
C. brandeiroi (Flach, 1906) |
|
C. championi (Reitter, 1902) |
|
C. cobosi (Báguena, 1955) |
|
C. dinizi (Branco, 1981) |
|
C. isabellae López-Colón, 1993 |
|
C. longitarsis (Iliger, 1803) |
|
C. luctuosa (Báguena, 1955) |
|
C. lusitanica (Baraud, 1975) |
|
C. mauritanica (Rambur, 1843) |
|
C. moelleri (Flach, 1906) |
|
C. stenostoma (Báguena, 1955) |
|
C. tangeriana (Kraatz, 1882) |
|
C. transtagana (Branco, 1981) |
|
C. zuzartei (Branco, 1981) |
|
• Elaphocera Gené, 1836 |
|
E. alonsoi López-Colón, 1992 |
|
E. ampla Báguena, 1955 |
|
E. angusta Kraatz, 1882 |
|
E. ardoini Baraud, 1966 |
|
E. autumnalis Motschulsky, 1859 |
|
E. baetica Sáez-Bolaño, 1993 |
|
E. baguenae Mancini, 1925 |
|
E. barbara Rambur, 1843 |
|
E. bicolorea Baraud, 1966 |
|
E. cacerensis (López-Colón & Rodr.-Arias, 1986) |
|
E. capdeboui Schauffus, 1882 |
|
E. carteiensis Rambur, 1843 |
|
E. christina Baraud, 1966 |
|
E. churianensis Rambur, 1843 |
|
E. cretica Reitter, 1902 |
|
E. elongata Schauffus, 1874 |
|
E. emarginata (Gyllenhal, 1817) |
|
E. erberi Kraatz, 1882 |
|
E. erichsoni Jaquelin du Val, 1860 |
|
E. ferreri (López-Colón & Plaza-Lama, 1989) |
|
E. gracilis Waltl, 1838 |
|
E. heydeni Kraatz, 1882 |
|
E. hiemalis Erichson, 1840 |
|
E. hirticollis Kraatz, 1882 |
|
E. hispalensis Rambur, 1843 |
|
E. ibicensis Escalera, 1926 |
|
E. insularis Fairmaire, 1877 |
|
E. lajonquierei Baraud, 1966 |
|
E. margaritae (Blanco-Villero, 1988) |
|
E. martini López-Colón, 1993 |
|
E. martorelli Fairmaire, 1879 |
|
E. nigroflabellata Burmeister, 1855 |
|
E. nupcialis Ruiz & Ávila, 1995 |
|
E. ochsi Baraud, 1987 |
|
E. pereplopezi Ruiz, 1996 |
|
E. phungae Baraud, 1987 |
|
E. segurensis (Escalera, 1923) |
|
E. staudingeri Reitter, 1902 |
|
E. sulcatula Fairmaire, 1884 |
|
E. suturalis Schauffuss, 1874 |
|
E. syriaca Kraatz, 1882 |
|
E. tethys Reitter, 1902 |
|
E. torressalai Báguena, 1955 |
|
• Europtron Marseul, 1867 |
|
E. gracile Marseul, 1867 |
|
E. confusum Marseul, 1878 |
|
• Hemictenius Reitter, 1902 |
|
H. gracilipes Semen, 1890 |
|
H. latitarsis Reitter, 1891 |
|
H. lebedevi Reitter, 1908 |
|
H. magnitarsis Reitter, 1897 |
|
H. nigrociliatus Reitter, 1897 |
|
H. nitidipes Reitter, 1902 |
|
H. opacipes Reitter, 1902 |
|
H. opacus Ball, 1870 |
|
H. ochripennis Reitter, 1902 |
|
H. simplicitarsis Reitter, 1897 |
|
H. tekkensis Reitter, 1897 |
|
• Kryzhanovskia Nikolaev & Kabakov, 1977 |
|
K. olegi Nikolaev & Kabakov, 1977 |
|
• Leptochristina Branco & Baraud, 1991 |
|
L. pubimargo (Reitter, 1902) |
|
L. pubicavula (Reitter, 1902) |
|
L. variabilis (Petrovitz, 1962) |
|
L. annamariae Branco & Baraud, 1991 |
|
• Otoclinius Brenske, 1896 |
|
O. fragilis Petrovitz, 1980 |
|
O. gracilipes Brenske, 1896 |
|
O. loebli Baraud, 1991 |
|
O. schauffelei Petrovitz, 1958 |
|
• Pachydema Castelnau, 1832 |
|
P. anthracina Fairmaire, 1860 |
|
P. bipartita (Brullé, 1838) |
|
P. carthaginensis Rambur, 1843 |
|
P. castanea (Brullé, 1838) |
|
P. fortunatorum Baraud, 1985 |
|
P. fuscipennis (Brullé, 1838) |
|
P. hornbecki Lucas, 1859 |
|
P. hirticollis (Fabricius, 1787) |
|
P. jeannei Baraud, 1985 |
|
P. lesnei Peyerimhoff, 1927 |
|
P. marmottani Fairmaire, 1868 |
|
P. menieri Baraud, 1985 |
|
P. nitidicollis Fairmaire, 1876 |
|
P. obscurata Fairmaire, 1883 |
|
P. obscura (Brullé, 1838) |
|
P. obscurella (Wollaston, 1864) |
|
P. palposa Reitter, 1898 |
|
P. rubripennis Lucas, 1848 |
|
P. tinerfensis Galante & Stebnicka, 1992 |
|
P. valdani Lucas, 1859 |
|
P. wollastoni 1927 |
|
P. xanthochroa Fairmaire, 1879 |
|
P. zhora Normand, 1951 |
|
• Pachydemocera Reitter, 1902 |
|
P. lucidicollis (Kraatz, 1882) |
|
• Peritryssus Reitter, 1918 |
|
P. excisus Reitter, 1918 |
|
• Pseudopachydema Balthasar, 1930 |
|
P. caucasica Balthasar, 1930 |
|
• Tanyproctoides Petrovitz, 1971 |
|
T. arabicus Petrovitz, 1971 |
|
• Tanyproctus Faldermann, 1835 |
|
T. bucharicus Reitter, 1897 |
|
T. confinis Motschulsky, 1859 |
|
T. eversmanni Reitter, 1902 |
|
T. ganglbaueri Brenske, 1897 |
|
T. iranicus Petrovitz, 1968 |
|
T. jordanicus Baraud, 1991 |
|
T. kindermanni Reiche, 1861 |
|
T. ovatus Motschulsky, 1859 |
|
T. pamphilus Petrovitz, 1967 |
|
T. persicus (Ménétries, 1832) |
|
T. rufidens Marseul. 1879 |
|
T. rugulosus Fairmaire, 1892 |
|
T. riparius Petrovitz, 1963 |
|
T. satanas Reitter, 1902 |
|
T. saulcyi Reiche, 1856 |
|
T. speculator Petrovitz, 1963 |
|
T. subciliatus Reitter, 1902 |
|
T. verryi Fairmaire, 1884 |
Metodología
 |
 |
 |
 |
 |
 |
 |
|
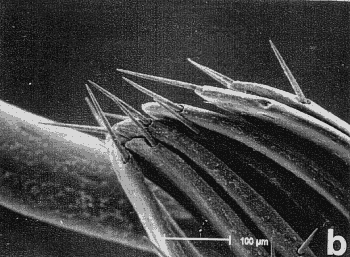
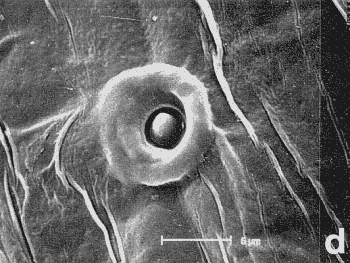
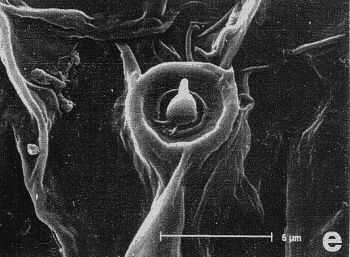
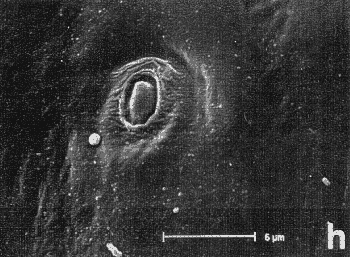
Figura 7. Micrografía electrónica de barrido del endofalo mostrando: sedas sensoriales de la superficie de la antena (a, b) y las sedas basicónicas (‘pegs’), situadas encima del extremo apical de la Pieza en ‘V’ (c, d, e, f y h), que aparecen en ciertos géneros de Pachydeminae tales como Pachydemocera, Ceramida y Elaphocera. |
En la búsqueda de nuevos rasgos de valor taxonómico que pudieran ser utilizados posteriormente en la reconstrucción filogenética, se exploraron cuatro tipos distintos de caracteres: morfología externa, aparato bucal (fig. 2), anatomía alar (fig. 3), la anatomía y morfología genital de ambos sexos: el tegmen y saco interno del edeago en los machos (figs. 4 y 5) y el sistema genital femenino (fig. 6). El estudio de estos caracteres implicó metodologías distintas pero ya bien establecidas en los estudios de morfología y anatomía comparada de insectos: observación directa de los ejemplares bajo lupa binocular, disección de piezas bucales y genitalia de ambos sexos (más de 600), preparaciones microscópicas, dibujo con cámara clara y videoimpresiones mediante cámara de vídeo conectada a monitor y estereomicroscopio. Para el análisis de estructuras de pequeño tamaño se empleó el microscopio electrónico de barrido ('Scanning') (fig. 7); no se utilizó el “punto crítico”, ya que se comprobó que con esta técnica, el endofalo se enrollaba a veces sobre su cara interna.
En el caso del género Ceramida, además de los caracteres morfológicos cualitativos se emplearon caracteres continuos (medidas) en la separación de las especies. La toma de medidas se realizó con un medidor digital de resolución 0.001 mm. Estas medidas fueron analizadas utilizando técnicas de análisis morfométrico multivariante: Análisis múltiple de la varianza (MANOVA), Análisis Discriminante y Análisis de Componentes Principales. En Ceramida los únicos estudios morfométricos, se han limitado a la discriminación de pares de especies empleando ciertas medidas (Serrano, 1985). El problema es que la separación morfométrica de las especies se ha evaluado mediante análisis univariante de cada carácter por separado. Este tipo de enfoque es erróneo puesto que asume que no existe correlación entre los caracteres y que éstos son independientes (Willig et al., 1986; Willig & Owen, 1987). Pero la respuesta de un organismo a las fuerzas selectivas no se manifiesta muchas veces como un cambio en un sólo carácter sino como la adaptación de un conjunto de caracteres morfológicos interrelacionados. Esto es especialmente importante en el caso de los caracteres continuos, fuertemente correlacionados como resultado de su integración genética: pleiotropía. Como el análisis univariante no tiene en cuenta esta integración genética, a menudo sobreestima las diferencias globales entre grupos y conduce a conclusiones sistemáticas erróneas (Willig et al., 1986). El análisis multivariante, en cambio, considera la intercorrelación de los caracteres al evaluar las diferencias intergrupales. Por tanto, es el método estadístico más apropiado para estudiar la taxonomía de grupos como Ceramida.
La reconstrucción filogenética se realizó de acuerdo con los principios teóricos y metodológicos del análisis cladista, bajo el supuesto de máxima parsimonia. Se utilizó como criterio de polaridad el grupo externo. Como tal, se consideraron cuatro especies representantes de cuatro linajes basales de Melolonthidae (Coca Abia, 1995). Se incluyeron caracteres binarios y caracteres multiestado, ordenados y desordenados. En todos los análisis el número de caracteres fue superior al número de taxones, 63 en el análisis de la subfamilia y 39 en la filogenia de los géneros Elaphocera y Cermida.
En la búsqueda de la reconstrucción filogenética más parsimoniosa, se emplearon dos programas de análisis filogenético: Hennig86 versión 1.5 (Farris, 1985) y Paup versión 3.1 (Swofford, 1991). Ambos están basados en el principio de máxima parsimonia pero difieren en los criterios de parsimonia de los algoritmos empleados en la reconstrucción filogenética. En la búsqueda de la reconstrucción filogenética más parsimoniosa no se pudo emplear la “búsqueda exhaustiva”, ya que el tamaño de las matrices de datos requería un tiempo de cálculo excesivo. En su lugar se emplearon dos técnicas distintas que reducen el tiempo de búsqueda: “branch and bound” (poda y reinjerto) y “búsqueda heurística”. Cuando el análisis filogenético proporcionó más de un árbol igualmente parsimonioso, se realizaron dos análisis diferentes: pesaje sucesivo (Farris, 1969) y la obtención de árboles de consenso; estos últimos se obtuvieron con el programa Component 2.0 (Page, 1993). Una vez obtenidos los árboles más parsimoniosos, se mapeó la distribución y los cambios entre estados de carácter dentro de cada árbol, mediante el programa Clados 1. 1. (Nixon, 1992). Clados efectúa los dos procesos de optimización de caracteres con los que trabajan Paup y Hennig86: Deltran (“delayed transformation optimization”) y Actran (“accelerated transformation optimization").
El análisis biogeográfico histórico se efectuó con la ayuda del programa Diva (Ronquist, 1996, 1997): "dispersion-vicariance analysis". Diva no restringe las relaciones entre áreas a un patrón cladogenético jerárquico, ni hace ninguna asunción a priori sobre la forma o la existencia de un único escenario biogeográfico. Por tanto, permite que las relaciones entre áreas sean reticuladas y múltiples, es decir, puede utilizarse en grupos afectados de distinta forma por sucesivos eventos geológicos. El resultado del análisis no es un cladograma de áreas, sino una reconstrucción de la evolución histórica de las áreas ancestrales del grupo estudiado. Por otro lado, en la reconstrucción de distribuciones ancestrales, Diva considera explícitamente los eventos de dispersión y extinción buscando la hipótesis biogeográfica más parsimoniosa, es decir, aquella en la que es necesario postular el menor número de dispersiones y extinciones para explicar la distribución geográfica actual.
Resumen de Resultados y Conclusiones
1. Exploración de Caracteres de Valor Taxonómico
El estudio de nuevas estructuras anatómicas, como el aparato bucal, la anatomía alar, o el sistema genital, proporcionaron nuevos caracteres de valor taxonómico. Estos caracteres presentan menor variabilidad interespecífica que los caracteres de morfología externa utilizados tradicionalmente. En particular, el aparato bucal (fig. 2) y la anatomía alar (fig. 3) proporcionan buenos caracteres en los rangos genéricos. El sistema genital proporciona caracteres de valor filogenético y la genitalia masculina permite, además, distinguir especies (figs. 3 y 4). La genitalia femenina, en cambio, tan sólo muestras rasgos sistemáticos en el rango de género (fig. 6)
2. Taxonomía
2.1. Género Elaphocera Gené, 1836:
La revisión taxonómica del género Elaphocera Gené, 1836 permitió reconocer 19 especies. Este nuevo catálogo supone una reducción importante del número de taxones incluidos hasta ahora en el género: veinticinco taxones pasan a ser nuevas sinonimias. Se describe además una nueva especie. De las 19 especies, diez son endemismos íbero-baleares (ver Apéndice) (50% de las especies), mientras que el resto colonizan Cerdeña (dos), el Mediterráneo oriental (cinco) y el Norte de África (dos especies). El elevado número de sinonimias se explica porque hasta hoy, carecíamos de un estudio comparado de todas las especies del género. Por otro lado, la obra de conjunto de Jacques Baraud (1992) introdujo numerosos errores al no estudiar el autor el material tipo de muchas especies. Estos errores fueron luego recogidos por otros autores contribuyendo, de esta forma, a incrementar la confusión taxonómica.
2.2. Género Ceramida Baraud, 1987
Como resultado de la revisión taxonómica, se reconocieron once especies dentro del género. Ocho taxones pasan a ser nuevas sinonimias. El estudio de caracteres cualitativos y el análisis morfométrico, confirman este resultado y proporcionan nuevos caracteres diagnósticos para separar las once especies.
Sin embargo, en ocho de estas once especies se encontraron individuos con caracteres en mosaico y morfotipos de morfología 'intermedia' en los que los rasgos diagnósticos no son concluyentes. La presencia de estos morfotipos intermedios, implica la posibilidad de fenómenos de hibridación entre especies o, al menos, la existencia de morfoclines de especies muy polimórficas representadas por poblaciones divergentes en vías de especiación, pero en las que todavía no se han fijado los caracteres morfológicos. En resumen, de las dieciocho especies tradicionalmente reconocidas dentro del género Ceramida, solo cuatro pueden ser calificadas de 'buenas especies' según el concepto biológico y filogenético de especie: mantienen el aislamiento reproductor y son diagnosticables, al menos, por un carácter (ver Apéndice).
3. Filogenia y Biogeografía
Las ideas básicas de la investigación sobre la evolución de los Pachydeminae en el área paleártica, pueden resumirse en los siguientes puntos:
1. Los Pachydeminae paleárticos constituyen un grupo monofilético definido por tres sinapomorfías: i) esternitos abdominales no soldados en la mitad del segmento; ii) el quinto y el sexto esternito están separados por una gruesa membrana; iii) parámeros no esclerotizados ventralmente. Otras novedades evolutivas son el ensanchamiento de los artejos tarsales del macho, con un cepillo de sedas en su cara inferior y las uñas hendidas en el ápice. Los caracteres referidos al aparato bucal (fig. 2) y a la genitalia masculina (interna y externa; figs. 4 y 5) constituyen las mejores hipótesis de sinapomorfías en la definición de las relaciones entre géneros. Estos caracteres presentan menor variabilidad que los caracteres de morfología externa utilizados tradicionalmente.
2. Se acepta el carácter monofilético de los géneros Pachydema y Hemictenius, otros, en cambio, como Leptochristina son parafiléticos. Tanyproctus y Otoclinius, no tienen base filogenética. Tampoco es muy consistente la posición y definición taxonómica de algunos géneros menores como Atanyproctus y otros monoespecíficos tales como: Kryzhanovskya, Psudopachydema, Tanyproctoides, Brenskiella, Alaia y Pachydemocera. Asimismo, algunas especies han de ser reubicadas; e.g.: Pachydema zhora Normand, 1951 y Elaphocera barbara Rambur, 1843. Esta última, es la especie hermana del género Ceramida y ambos representan a su vez el grupo hermano del género Elaphocera. Estos dos géneros ibéricos son también monofiléticos.
3. El origen de los Pachydeminae está ligado a la zona occidental del paleocontinente Gondwana. Los linajes más antiguos de posición basal en el cladograma, todavía subsisten en el África paleártica y el grupo hermano de la subfamilia (Perytrissus) en Sicilia. Se trata por tanto, de un grupo muy primitivo cuyo origen es difícil precisar en el amplio lapso de tiempo que trascurre entre el Cretácico superior y el Oligoceno inferior.
4. Las primeras vicarianzas dividieron la fauna africana (género Pachydema) de los linajes asiáticos (Mioceno medio) y luego la fauna asiática en dos áreas: una occidental (Asia Occidental) y otra más oriental (Asia Central: llanura aralo-cáspica) en la que se diferenció el género Hemictenius (Mioceno superior).
5. Los presencia de los clados más derivados en tierras laurásicas, Península Ibérica y los Balcanes, se debió a dispersiones posteriores.
6. Los géneros ibéricos Elaphocera y Ceramida se originaron a partir de un linaje ancestral presente en Asia Menor que, al menos en dos ocasiones, se dispersó hacia el oeste por vía meridional (norteafricana). Estas dispersiones dieron lugar a sendas vicarianzas este-oeste que separaron primero al género Ceramida de Elaphocera y luego a éste último, en dos linajes: oriental y occidental (Mioceno superior-Plioceno). Posteriormente estos géneros especiaron repetidamente, pero esta especiación ha sido menor que la que sugieren los estudios taxonómicos anteriores a nuestra revisión.
7. La presencia de Pachydeminae en la Península Ibérica es una condición biogeográfica derivada, resultado de la dispersión de linajes transmediterráneos de origen asiático, a través del África paleártica. Este patrón coincide aproximadamente con el cladograma de áreas propuesto recientemente por De Jong (1998) para otros grupos mediterráneos pero en cambio, contradice la hipótesis de Oosterbroek & Artnzen (1992) según la cual la Península Ibérica representa el área plesiocórica (primitiva) para los grupos de distribución mediterránea.
Literatura Citada
Alvarado, M., Serrano, A., De la Rosa, A. & Durán, J. M., 1995. Problemática de los gusanos blancos en el olivar de la provincia de Sevilla. V Jornadas Cinetíficas de la Sociedad Española de Entomología Aplicada, Sevilla, 1995.
Alvarado, M., Serrano, A., Durán, J. M. & De la Rosa, A., 1996. Problemática de los gusanos blancos (Coleoptera, Scarabaeidae) en el olivar de la provincia de Sevilla. Boletín de Sanidad Vegetal, Plagas, 22: 319-328.
Báguena, L., 1955. Observaciones sobre las especies ibéricas de Elaphocera Gené. Eos, 31 (1-2): 123-153.
Balachowsky, A. S., 1962. Entomologie appliquée à l’Agriculture, vol. 1º: Coléoptères. Masson et Cie., Paris: 564 pp.
Balleto, E. & Casale, A., 1991. Mediterranean Insect Conservation. En: The Conservation of Insects and their Habitats. pp: 121-140. Collins, N. M. & Thomas, J. A. eds., Academic Press. 1991.
Baraud, J., 1985. Coléoptères Scarabaeoidea. Faune du Nord de l’Afrique: du Maroc au Sinaï. Encyclopédie Entomologique, XLVI. Lechevalier. 651 pp.
Baraud, J., 1991. Otoclinius loebli, nouvelle espèce d'Iran (Coleoptera, Melolonthidae, Pachydeminae). Arschs. Sci Genéve, 44 (3): 289-291.
Baraud, J., 1992. Coléoptères Scarabaeoidea d’Europe. Faune de France et régions limitrophes, 78. Société Linnéenne de Lyon et Fédération Francaise des Sociétés de Sciences Naturelles. Lyon. 850 pp.
Baraud, J. & Branco, T.,1991. Scission du genre Pachydemocera Reitter, 1902 (Coleoptera, Melolonthidae, Pachydeminae). Bulletin de la Société entomologique de France, 96 (2): 175-181.
Blanco-Villero, J. M., 1988. Elaphocerida margaritae, nueva especie de la Península Ibérica (Coleoptera, Melolonthidae). Novelle Revue Entomologie (N.S.), 5 (3): 238-286.
Branco, T., 1981. Contribution à la connaisance des Elaphocera Gené ibériques: quatre nouvelles espèces du Portugal (Col. Scarabaeoidea, Melolonthidae). Bulletin de la Société Entomologique de France, 86, 124-144.
Coca Abia, M.,1995. Taxonomía, Filogenia y Biogeografía del género Rhizotrogus (Col. Melolonthidae) en el Mediterráneo occidental. Tesis Doctoral (inédita). UCM. Madrid. 234 pp.
De Jon, H., 1998. In search of historical biogeographic patterns in the Western mediterranean terrestrial fauna. Biological journal of the Linnean Society, 65: 99-164.
Edelstam, C., Skoog, G. Lundberg, H. & Ramel, C., 1992. Perspective in the maintenace of Biodiversity. Report. Royal Academy of Sciences, 30 p.
Farris, J. S., 1969. A succesive approximations approach to character weighting. Systematic Zoology, 18: 374-385.
Farris, J. S., 1988. HENNIG86 version 1.5; sofware and MSDOS program.
López-Colón, J. I., 1986. Elaphocerida cacerensis, nueva especie extremeña (Coleoptera, Melolonthidae). Nouvelle Revue Entomologie (N. S.), 3 (1): 99-101.
Machado, A., 1998. Biodiversidad. Un paseo por el concepto y las Islas Canarias. Ed. Cabildo insuar de Tenerife. Sanata Cruz de Tenerife. 67 pp.
Margalef, R.,1997. Our Biosfere. En: Excellence in Ecology, 10 (ed. O. Kinner). Ecology Institute, Oldendorf, 176 pp.
Martín Piera, F., 1997. Apuntes sobre Biodiversidad y Conservación de Insectos: Dilemas, Ficciones y ¿Soluciones?. Boletín de la Sociedad entomológica aragonesa, 20: 25-55.
Medvedev, S. I., 1960. Description of larvae of eight Scarabaeid species from the Ukraine and Central Asia. Zoologichesky Zhurnal, 39: 381-393.
Nikolaev, G. V., 1987. Scarabaeoidea (Coleoptera) of Kazakhstan and Central Asia. Nauka, Alma Ata.
Nixon, K. C., 1993. CLADOS. version 1.4. Computer software and documentation. published by the author. P. O. Box 270, Tumansburg, NY, 14886 USA.
Oosterbroeck, P. & Arntzen, J. W., 1992. Area-Cladograms of circummedite-rranean taxa in relation to Mediterranean paleogeography. Journal of Biogeography, 19: 3-20
Page, R. D. M., 1993. 'Component'. Users manual and software. Release 2.0. Published by the author.
Piatella, E. & Sabatinelli, G., 1996. Una nuova specie di Tanyproctus delle Sporadi meridionale. Fragmenta entomologica, 27 (2): 361-368.
Ronquist, F., 1996. 'Diva', version 1.1. User’s Manual. Computer program for MacOS and Win32. Available from www.systbot.uu.se/ personel/ f_ronquist.html.
Ronquist, F., 1997. Dispersal-Vicariance analysis : a new biogeographic approach to the quantification of historical biogeography. Systematic Biology, 46(1) : 195-203.
Ruiz, J. L. & Ávila, J. M., 1995. Elaphocera nupcialis n. sp. del Sureste de la Península Ibérica (Coleoptera, Scarabaeoidea: Melolonthinae). Boletín de la Asociación española de Entomología, 19 (1-2): 243-251.
Serrano, A. R. M.., 1985.-Contribution à l’étude du genre Elaphocera Gené, 1836 en Algarve (sud de Portugal). Description de deux espèces nouvelles (Coleoptera, Melolonthidae). Nouvelle Revue de Entomologie (N. S.): 2 (4), 359-370.
Serrano, A., Alvarado, M., Durán, J. M. & De la Rosa, A., 1996. Contribución al conocimiento de Ceramida (Elaphocera) spp. (Coleoptera: Scarabaeidae) plaga de los olivares de la provincia de Sevilla. Boletín de Sanidad Vegetal, Plagas, 22: 203-211.
Swofford, D. L., 1991. 'Paup': Phylogenetic analysis using parsimony, version 3. 1. Illinois Natural History Survey, Champaign, Illinois.
Willig, M.R. & Owen, R.D., 1987.- Univariate analyses of morphometric variation do not emulate the results of multivariate analyses. Systematc Zoology, 36 (4): 398-400.
Willig, M.R., Owen, R.D. & Colbert, R.L., 1986.- Assesment of morphometric variation in natural populations: the inadequacy of the univariate approach. Systematic Zoology, 35 (2), 195-203.
Página creada en diciembre de 2001
Última actualización: lunes, 31 de diciembre de 2001